Zhenli Huang* and Haiying Li
Sci. Adv. 10, eadi6580 (2024) Submitted 10 May 2023
Accepted 5 April 2024
Published 10 May 2024
China Institute of Water Resources and Hydropower Research, Beijing 100038,China.
中国水利水电科学研究院,北京 通讯作者:Corresponding author. Email: zhlhuang@263.net
The impact of dams on global migratory fish stocks is a major challenge and remains seriously underestimated.
大坝对全球迁徙性鱼类种群的影响是一个重大挑战,并且至今被严重低估。
China has initiated a dozen fish rescue programs for the dams on the Yangtze River, focusing on five flagship species—Chinese sturgeon, Chinese paddlefish, Yangtze sturgeon, Chinese sucker, and Coreius guichenoti.
中国已在长江上的大坝启动了十几个鱼类救援计划,重点关注五种旗舰物种——中华鲟、白鲟、长江鲟、中华鲴和鯮。
Despite 40 years of effort, these five fishes are on the verge of extinction.
尽管经过40年的努力,这五种鱼类仍濒临灭绝。
Here, we propose an analytical tool that includes a framework of fish migration taxonomy and six life cycle models, the concepts of invalid stock and the dam impact coefficient, and a simplified population model.
在这里,我们提出了一个分析工具,包括一个鱼类迁徙分类框架和六个生命周期模型,无效种群和大坝影响系数的概念,以及一个简化的种群模型。
We then clarify the migration patterns and life cycles of these fishes and show that the Yangtze dams have severely disrupted the life cycle integrity of these species, causing seven types of invalid stocks and their exponential population declines.
然后,我们阐明了这些鱼类的迁徙模式和生命周期,并展示了长江大坝严重破坏了这些物种的生命周期完整性,导致七种无效种群及其指数级种群下降。
Last, we discuss six scientific misjudgments underpinning the fish rescue programs and recommend reforms to China’s fish rescue strategy.
最后,我们讨论了支撑鱼类救援计划的六个科学误判,并推荐了中国鱼类救援策略的改革。
INTRODUCTION 引言
Dams have engendered major challenges for migratory fish globally (1–4).
大坝在全球范围内为迁徙性鱼类带来了重大挑战(1-4)。
More than 58,000 dams higher than 15 m have been built in most of the world’s rivers (5) because most countries have overwhelmingly focused on building dams for the multi-objective utilization of freshwater resources themselves while ignoring or overlooking the huge economic and ecological value of migratory fish.
世界上大多数河流上已建造了超过58,000座高度超过15米的大坝(5),因为大多数国家在利用淡水资源方面有着压倒性的关注,同时忽视或忽略了迁徙性鱼类的巨大经济和生态价值。
In February 2021, 16 conservation organizations released a report, The World’s Forgotten Fishes, declaring that migratory fish stocks have declined by 76% since 1970, and worldwide populations of “mega fish” have declined by an astounding 94%.
2021年2月,16个保护组织发布了一份报告,《世界被遗忘的鱼类》,宣布自1970年以来,迁徙性鱼类种群下降了76%,而“巨型鱼类”的全球种群数量惊人地下降了94%。
Globally, threats to migratory fish have been linked to factors other than dams, including overfishing, water pollution, and sand mining (6).
全球而言,对迁徙性鱼类的威胁已与大坝以外的因素联系起来,包括过度捕捞、水污染和采砂(6)。
Because of a lack of applicable tools and available data, it has been impossible to distinguish the relative contributions of dam and non-dam factors, leading to large discrepancies between expected results and reality and, crucially, a pervasive underestimation of dam impacts (7).
由于缺乏适用的工具和可用数据,无法区分大坝和非大坝因素的相对贡献,导致预期结果与现实之间存在巨大差异,关键是普遍低估了大坝的影响(7)。
Today, the recovery of flagship migratory species from population collapse or extinction in large dammed rivers is becoming an intractable global problem.
今天,在大型水坝河流中,旗舰迁徙性物种从种群崩溃或灭绝中恢复成为一个棘手的全球问题。
This includes the various sturgeon species affected by the Iron Gates I/II on the Danube River in Europe, the Volgograd Dam on the Volga River in Russia, and the Three Gorges Dam (TGD) on the Yangtze River in China (8–10).
这包括受欧洲多瑙河上的铁门I/II大坝影响的各种鲟鱼物种,俄罗斯伏尔加河上的伏尔加格勒大坝,以及中国长江上的三峡大坝(TGD)(8-10)。
The Yangtze River, which is the longest river in China and the third longest in the world, spans 6387 km with a drainage area of 1.8 million km2 (11).
长江是中国最长、世界第三长的河流,全长6387公里,流域面积180万平方公里(11)。
Originating from the snow- covered mountain Geladandong on the Qinghai- Tibet Plateau, it is divided into three reaches: the upper (4504 km) above Yichang, the middle (955 km) from Yichang to Jiujiang, and the lower (938 km) from Jiujiang to the river mouth.
它起源于青藏高原上覆盖着雪的格拉萨东山,分为上游(4504公里)在宜昌以上,中游(955公里)从宜昌到九江,下游(938公里)从九江到河口。
The stretch of the river from Zhimenda to Yibin is known as the Jinsha River (3464 km).
从志门达到宜宾的河流段被称为金沙江(3464公里)。
It is divided into three sections: the upper from Zhimenda to Shigu, the middle from Shigu to Panzhihua, and the lower from Panzhihua to Yibin.
它分为三个部分:上游从志门达到石鼓,中游从石鼓到攀枝花,下游从攀枝花到宜宾。
The Yangtze River is China’s most abundant water resource, with the highest freshwater fish biodiversity and the largest number of migratory fish species.
长江是中国最丰富的水资源,拥有最高的淡水鱼类生物多样性和最多的迁徙性鱼类物种。
It is also home to more than 400 species of fish, 22 of which are listed as nationally protected species (12).
它还是400多种鱼类的家园,其中22种被列为国家保护物种(12)。
Hydropower development on the mainstem of the Yangtze River began in the 1970s with the Gezhouba Dam (GD).
长江干流的水力发电开发始于20世纪70年代的葛洲坝(GD)。
By the end of 2021, a dozen large dams had been successfully constructed on the upper Yangtze and middle/lower Jinsha rivers (Fig. 1, text S1, and table S1), with more under construction on the upper Jinsha.
截至2021年底,已在长江上游和金沙江中下游成功建成了十几座大型水坝(图1,文本S1和表S1),金沙江上游还有更多的水坝正在建设中。
These dams form China’s largest hydropower base and contribute to improving flood control, navigation, and power supply.
这些大坝构成了中国最大的水电基地,有助于改善防洪、航运和电力供应。
When the GD, the first dam across the mainstream of the Yangtze River, was built in the 1970s, the Chinese government explicitly demanded that the dam consider the conservation of fish.
当GD,长江主流上的第一座大坝在20世纪70年代建成时,中国政府明确要求大坝考虑鱼类保护。
To this end, a heated debate over conservation targets and fish passages raged from 1981 to 1982 (13).
为此,从1981年到1982年展开了关于保护目标和鱼道的激烈辩论(13)。
In late 1982, the Chinese government adopted the suggestions of the Institute of Hydrobiology (IHB) of the Chinese Academy of Sciences that the Chinese sturgeon (Acipenser sinensis) should be the sole focus of conservation efforts (14).
1982年底,中国政府采纳了中国科学院水生生物研究所(IHB)的建议,将中华鲟(Acipenser sinensis)作为保护工作的唯一重点(14)。
Subsequently, the IHB conducted nearly all of the studies on fish conservation in the 12 cascade dams (15, 16), identifying five fish species as key targets for conservation: the Chinese sturgeon, the Chinese paddlefish (Psephurus gladius), the Yangtze sturgeon (Dabry’s sturgeon, Acipenser dabryanus), the Chinese sucker (Myxocyprinus asiaticus), and Coreius guichenoti.
随后,IHB几乎进行了所有关于12级联大坝的鱼类保护研究(15, 16),确定了五种鱼类作为保护的关键目标:中华鲟、中华匙吻鲟(Psephurus gladius)、长江鲟(达氏鲟,Acipenser dabryanus)、中华鲴(Myxocyprinus asiaticus)和鯮(Coreius guichenoti)。
The primary conservation measures for each dam—including restocking, fishing bans, and fish natural reserves—did not include targeted interventions, such as the construction of fishways to promote upstream and downstream connectivity or environmental flows to mitigate hydrological shifts.
每个大坝的主要保护措施——包括人工增殖放流、禁渔令和鱼类自然保护区——没有包括针对性的干预措施,如建造鱼道以促进上下游连通性或环境流量以减轻水文变化的影响。
After four decades of implementation, the conservation efforts for each dam, collectively known as the Fish Rescue Programs (FRPs) of the Yangtze dams, have failed to achieve their goal of protecting the five flagship fishes (17–20).
经过四十年的实施,每个大坝的保护工作,统称为长江大坝的鱼类救援计划(FRPs),未能实现保护五种旗舰鱼类的目标(17-20)。
However, the failure of the FRPs provides a rare opportunity to examine the true impact of dams on migratory fish.
然而,FRPs的失败为检验大坝对迁徙性鱼类真正影响提供了一个难得的机会。
Dams can harm migratory fish by disrupting their life cycles and then causing population extinctions.
大坝可以通过扰乱它们的生命周期,然后导致种群灭绝来伤害迁徙性鱼类。
However, the quantitative effects of dams on these species are poorly understood, causing a substantial underestimation of their overall impact.
然而,大坝对这些物种的数量效应了解甚少,导致对它们整体影响的严重低估。
This is due to the lack of a clear classification system for fish migration, particularly for potamodromous fish, which results in a skewed understanding of the various migration patterns and corresponding life cycles.
这是由于缺乏明确的鱼类迁徙分类系统,特别是对于河川洄游性鱼类,导致对各种迁徙模式和相应生命周期的偏颇理解。
Moreover, the lack of standardized methodologies for identifying complete life cycles has resulted in a notable scarcity of global-scale life cycle datasets (21).
此外,缺乏标准化方法来识别完整的生命周期,导致全球尺度生命周期数据集的显著匮乏(21)。
In addition, the inability to conduct a quantitative evaluation of the majority of migratory fish due to data limitations exacerbates this issue.
此外,由于数据限制,无法对大多数迁徙性鱼类进行定量评估,加剧了这一问题
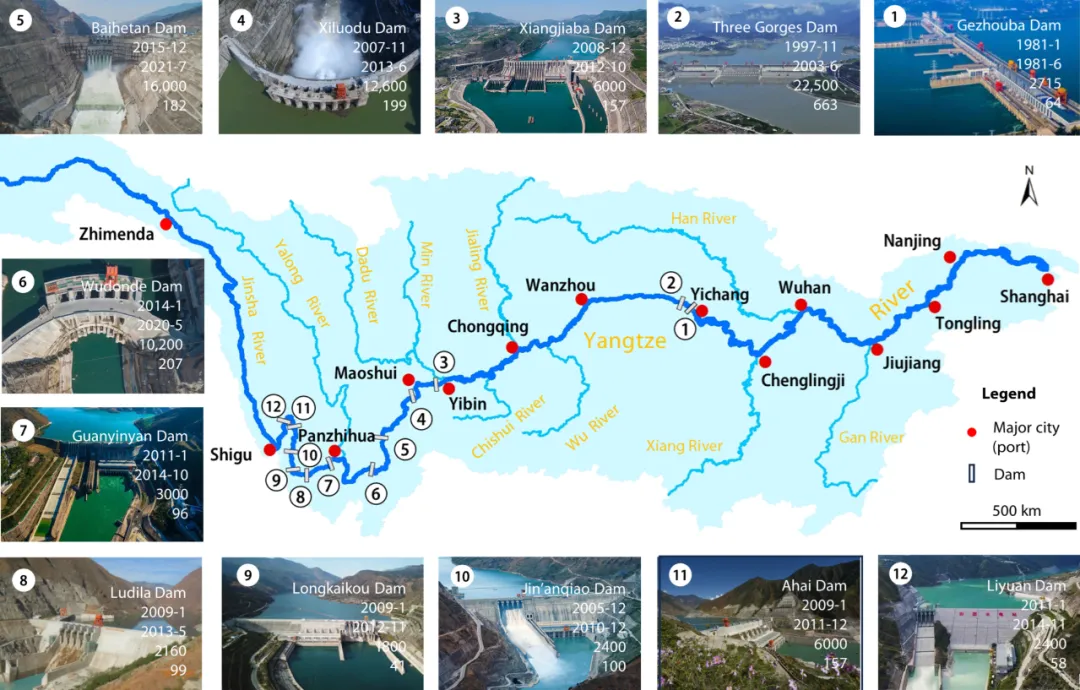
Fig. 1. Major cascade dams in the mainstream of the Yangtze River. By the end of 2021, a dozen large-scale cascade dams had been completed in the upper reaches of the Yangtze River and the middle and lower sections of the Jinsha River. The lowermost of these is the Gezhouba dam, preceded upriver by the Three Gorges Dam (TGD), Xiangjiaba Dam, Xiluodu Dam, Baihetan Dam, Wudongde Dam, Guanyinyan Dam, Ludila Dam, Longkaikou Dam, Jin’anqiao Dam, Ahai Dam, and Liyuan Dam. The inner information presented in the images of the 12 dams includes the dam’s name, river closure time (year–month), impoundment time (year–month), installed capacity [megawatt (MW)], and backwater length (km), arranged from top to bottom. All photo credits: Zhenli Huang and Haiying Li.
图1. 长江主流的主要梯级大坝。截至2021年底,长江上游和金沙江中下游已完成了十几座大型梯级大坝。这些大坝中最下游的是葛洲坝,上游依次是三峡大坝(TGD)、向家坝大坝、溪洛渡大坝、白鹤滩大坝、乌东德大坝、观音岩大坝、鲁地拉大坝、龙开口大坝、金安桥大坝、阿海大坝和梨园大坝。12座大坝的图片中展示的内部信息包括大坝名称、截流时间(年-月)、蓄水时间(年-月)、装机容量[兆瓦(MW)]和回水长度(公里),按从上到下的顺序排列。所有照片由黄镇利和李海英提供。
Hence, we propose an analytical tool aimed at quantitatively evaluating and contrasting the effects of the Yangtze cascade dams on the five fish species, using them as a case study.
因此,我们提出了一个分析工具,旨在定量评估和对比长江梯级大坝对这五种鱼类的影响,以它们作为案例研究。
This tool includes a three-tiered migration classification system, six descriptive life cycle models, the concepts of invalid stocks and the dam impact coefficient (DIC), and a simplified population model.
该工具包括一个三级迁移分类系统、六个描述性生命周期模型、无效种群和大坝影响系数(DIC)的概念,以及一个简化的种群模型。
Here, we first determine the migration pattern and life cycle of each of the five fish species using the migration classification system and the life cycle model.
在这里,我们首先使用迁移分类系统和生命周期模型,确定这五种鱼类各自的迁移模式和生命周期。
Then, we analyze the barrier effect of dams on the life cycle of each species and reveal the underlying mechanisms contributing to the decline of fish populations by estimating the invalid stocks and DICs.
然后,我们分析大坝对每种物种生命周期的阻碍效应,并通过对无效种群和DIC的估计,揭示导致鱼类种群下降的潜在机制。
Consequently, we use the simplified model to reconstruct the population decline processes of these five fishes.
因此,我们使用简化模型重构这五种鱼类的种群下降过程。
Last, we discuss scientific misjudgments and recommend reforms to China’s dam-related fish rescue strategy.
最后,我们讨论了科学判断的失误,并为中国的大坝相关鱼类救援策略提出了改革建议。
RESULTS Identifying migration patterns and life cycles of the five fish species When designing dams, accurately understanding the migratory behavior and life cycle of fish is the foundation of fish conservation.
结果:识别五种鱼类的迁徙模式和生命周期。在设计大坝时,准确理解鱼类的迁徙行为和生命周期是鱼类保护的基础。
Without a scientific framework of migration classification and a life cycle analysis tool, incorrect assumptions and misjudgments may be made about migratory fish traits and behaviors.
没有迁徙分类的科学框架和生命周期分析工具,可能会对迁徙性鱼类的特征和行为做出错误的假设和判断。
To date, Chinese ichthyologists have typically used an outdated classification system of fish migration, dividing freshwater fish into “migratory,” “semi- migratory,” and “sedentary” (22).
迄今为止,中国的鱼类学家通常使用一个过时的鱼类迁徙分类系统,将淡水鱼划分为“迁徙性”、“半迁徙性”和“定居性”(22)。
As a result, the migration patterns and life cycles of migratory fish in China are typically unclear or incorrect (23–27), leading to the FRPs having a weak scientific foundation (11–16).
结果,中国迁徙性鱼类的迁徙模式和生命周期通常是不清晰或错误的(23-27),导致鱼类救援计划(FRPs)的科学基础薄弱(11-16)。
For example, the Chinese paddlefish has been described as “one of the world’s largest freshwater fishes” (17, 23), which overlooks its diadromous nature, and the migratory Yangtze sturgeon has been described as a “resident” freshwater fish (24) that does not migrate long distances (25), although its migratory distance is up to 2200 km.
例如,中华匙吻鲟被描述为“世界上最大的淡水鱼之一”(17, 23),这忽略了它的洄游性质,而迁徙性长江鲟被描述为一种“定居性”淡水鱼(24),它不会进行长距离迁徙(25),尽管它的迁徙距离可达2200公里。
There is an urgent need to reclassify the migratory fish of the Yangtze River and clarify their life cycles.
迫切需要对长江的迁徙性鱼类进行重新分类并阐明它们的生命周期。
Here, we propose a three- tiered and three- category framework for fish migration taxonomy, including definitions of various migration patterns, accompanied by six descriptive life cycle models of dam- related migration patterns (Fig. 2 and Methods).
在这里,我们提出了一个三级和三类框架的鱼类迁徙分类法,包括各种迁徙模式的定义,并伴随着六个描述性生命周期模型,这些模型涉及与大坝相关的迁徙模式(图2和方法)。
Within this framework, we further distinguish potamodromy into two types—riverine potamodromy and lacustrine potamodromy—with definitions and corresponding life cycle models.
在此框架内,我们进一步将河川洄游性鱼类分为两种类型——河流性河川洄游性和湖泊性河川洄游性——并提供定义和相应的生命周期模型。
Using a procedure for determining migratory fish life cycle (Methods), we can access the full life cycles of data-limited fish by identifying their migration patterns.
通过使用确定迁徙性鱼类生命周期的程序(方法),我们可以通过识别它们的迁徙模式来了解数据有限的鱼类的完整生命周期。
Our findings indicated that the migration patterns of the five fishes belonged to one of two categories—diadromy and potamodromy (Fig. 3 and text S2).
我们的发现表明,这五种鱼类的迁徙模式属于两大类之一——洄游性和河川洄游性(图3和文本S2)。
The Chinese sturgeon exhibits anadromy (type) of diadromy (category), which inhabits a marine environment for most of its life, swimming the 2850 km up to breed in the upper Yangtze River (Fig. 3A).
中华鲟展示了洄游性(类别)中的溯河洄游性(类型),它大部分生命时间生活在海洋环境中,游上2850公里到长江上游繁殖(图3A)。
The Chinese paddlefish exhibits the freshwater amphidromous subtype of the amphidromous type in the diadromous category, and it breeds in the upper Yangtze River; their juveniles need to migrate downriver to the sea, a distance of 2750 km.
中华匙吻鲟展示了洄游性类别中淡水两栖洄游亚型的两栖洄游性,它在长江上游繁殖;其幼鱼需要顺流而下迁移到海里,距离为2750公里。
The juveniles then undergo osmoregulation in saltwater in their early growth stage and then return to the Yangtze River (Fig. 3B).
然后,幼鱼在它们早期生长阶段在咸水中进行渗透压调节,然后返回长江(图3B)。
Because of this portion of its life cycle, the Chinese paddlefish cannot be considered a strictly freshwater (17, 23), anadromous (18), or potamodromous fish (28).
由于其生命周期的这一部分,中华匙吻鲟不能被视为严格的淡水鱼(17, 23)、洄游性鱼类(18)或河川洄游性鱼类(28)。
The remaining three species (the Yangtze sturgeon, the Chinese sucker, and C. guichenoti) exhibit the riverine type of potamodromy (category), breeding in the upper Yangtze River and completing their life cycles within freshwater, and their farthest migration distances are up to 2200, 2800, and 1700 km, respectively (Fig. 3, C to E).
剩下的三个物种(长江鲟、中华鲴和鯮)展示了河川洄游性(类别)的河流性类型,它们在长江上游繁殖并在淡水中完成生命周期,它们最远的迁徙距离分别可达2200公里、2800公里和1700公里(图3,C至E)。
We found no evidence of partial migration behavior (that is, populations composed of resident and migratory contingents) in any of the five fish species.
我们没有发现这五种鱼类中有任何部分迁徙行为的证据(即,由定居种群和迁徙群体组成的种群)。
We found that the life cycles of migratory fish in rivers usually include at least three bidirectional migrations for varying purposes at specific growth stages: the downriver migration of larvae and
我们发现,河流中迁徙性鱼类的生命周期通常包括至少三次双向迁徙,用于特定生长阶段的不同目的:幼体和
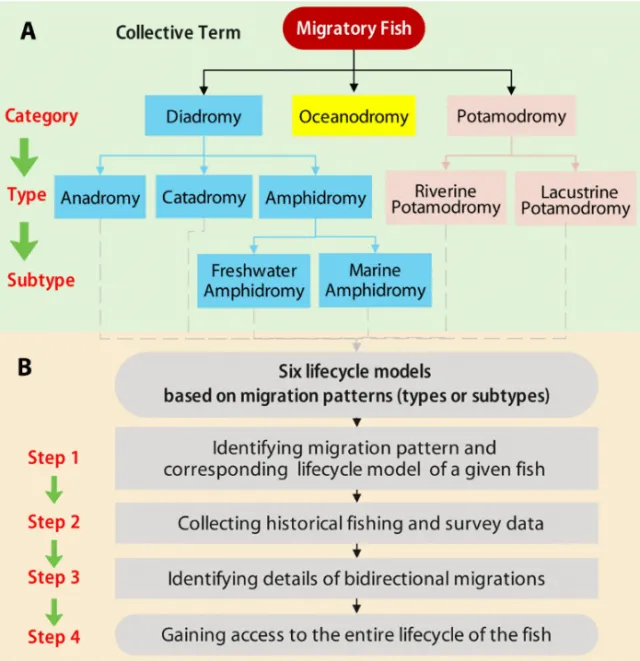
Fig. 2. Three-tiered framework of fish migration taxonomy and six descriptive life cycle models of migratory fish, which are related to dams. Here, we present the fish migration classification system (A), which introduces two types of potamodromy. On the basis of this system, we propose six descriptive life cycle models (B) that assist in acquiring the entire life cycles of data-limited fish species in four steps. See Methods and text S2 for definitions of migration types or subtypes and their corresponding life cycle models.
图2. 三层框架的鱼类迁徙分类和六种描述性生命周期模型,与大坝相关的迁徙性鱼类。这里,我们展示了鱼类迁徙分类系统(A),它引入了两种河川洄游性。基于这个系统,我们提出了六种描述性生命周期模型(B),这些模型帮助我们通过四个步骤获得数据有限的鱼类物种的整个生命周期。有关迁徙类型或亚型的定义及其相应的生命周期模型,请参见方法和文本S2
juveniles for feeding and refuge, the upriver migration of subadults and adults for reproduction, and the downriver migration of post-spawners for feeding.
幼鱼和少年鱼为了觅食和避难而进行的顺河迁移,亚成体和成体为了繁殖而进行的逆河迁移,以及产卵后个体为了觅食而进行的顺河迁移。
While we revealed that migration triggers are endogenous and linked to gonadal development (fig. S1), they may also be exogenous and depend on environmental factors, such as water temperature and fluctuations in water level.
虽然我们揭示了迁徙触发因素是内源性的,并且与性腺发育有关(图S1),但它们也可能来自外部,依赖于环境因素,如水温度和水位波动。
Next, we present the life cycle findings for each of the five fish species before the construction of the Yangtze dams.
接下来,我们将介绍在长江大坝建设之前,这五种鱼类生命周期的发现。
Chinese sturgeon
中华鲟
The Chinese sturgeon belongs to the order Acipenseriformes, the family Acipenseridae, and the genus Acipenser in the binomial system.
中华鲟属于二名法系统中的鲟形目、鲟科和鲟属。
It was granted first-class national protected animal status in China in 1988, listed as critically endangered (CR) on the International Union for Conservation of Nature’s Red List of Threatened Species (IUCN Red List) in 2010, and listed in CITES Appendix II in 1998 (29).
它在1988年被中国列为一级国家保护动物,在2010年被国际自然保护联盟濒危物种红色名录(IUCN红色名录)列为极危(CR),并在1998年被列入《濒危野生动植物种国际贸易公约》附录II(29)。
There are 19 spawning sites along an 800-km stretch of the Maoshui-Wanzhou section, with the major sites between Yibin and Hejiang, such as Sankuaishi (Fig. 3A), Pianyanzi, Jinduizi, Tielutan, and Wanglongqi (25).
在茅水-万州段800公里的范围内有19个产卵场,主要场地位于宜宾和合江之间,如三块石(图3A)、偏岩子、金堆子、铁炉滩和王龙溪(25)。
Breeding occurs during the last flood in October, commonly referred to as the “autumn water descending” phenomenon.
繁殖发生在10月的最后一次洪水期间,通常称为“秋水下降”现象。
Concurrently, water temperature decreases to 18° to 20°C, accompanied by an increase in flow velocity and sediment content.
同时,水温下降到18°至20°C,伴随着流速的增加和泥沙含量的上升。
Females reproduce between 13 and 34 years old and males between 8 and 27 years old, and the sex ratio (females to males) is 1:1.
雌性在13到34岁之间繁殖,雄性在8到27岁之间繁殖,性别比(雌性对雄性)为1:1。
The species migrate between Yibin and the sea, spending most of its life on the East China Sea’s continental shelf.
该物种在宜宾和大海之间迁徙,在大部分生命时间里生活在东海大陆架上。
The anadromous nature of the Chinese sturgeon has consistently remained undisputed.
中华鲟的溯河洄游性一直未受争议。
Nonetheless, the preceding depiction of its life cycle was overly simplistic and devoid of comprehensive migratory details.
然而,对其生命周期的先前描述过于简单,缺乏全面的迁徙细节。
In the case of this extensively examined species, we have acquired an intricate portrayal of its full life cycle within the Yangtze River using a migration dynamics model (text S4.1) (30, 31).
对于这个经过广泛研究的物种,我们使用迁徙动态模型(文本S4.1)获得了它在长江内完整生命周期的复杂描述(30, 31)。
As shown in Fig. 4A, adults at the gonadal stage III migrate to the Yangtze estuary every summer from June to August.
如图4A所示,处于性腺发育第三期的成体每年夏天从6月到8月迁移到长江口。
They then move up to the Hubei section in September, Yichang in December, and Yibin in July of the following year.
然后它们在9月迁移到湖北段,在12月到达宜昌,并在次年7月到达宜宾。
Once they reach Yibin Maoshui, a series of upstream rapids prevent them from moving further, so they congregate between Maoshui and Hejiang from July to October.
一旦它们到达宜宾茅水,一系列的上游急流阻止了它们进一步移动,因此它们从7月到10月聚集在茅水和合江之间。
Approximately 70% of the spawners complete their first breeding batch by mid-October, while the remaining 30% complete their second breeding batch before mid-November.
大约70%的产卵者在10月中旬完成第一批繁殖,而剩下的30%在11月中旬之前完成第二批繁殖。
After spawning, the post-spawners at gonadal stage II migrate downstream and reach the estuary within 16–28 days.
产卵后,处于性腺发育第二期的个体顺流而下,16至28天内到达河口。
After spawning at the Sankuaishi spawning site (an example), fertilized eggs sink and adhere to the gravel substrate and hatch after 5 days.
在三块石产卵场(一个例子)产卵后,受精卵下沉并附着在砾石基质上,5天后孵化。
The number of days post-hatching (dph) was used to characterize the age of individual fish.
孵化后天数(dph)被用来描述个体鱼的年龄。
The downstream migration of juveniles can be divided into three phases: drift stage (1 to 8 dph), cover stage (9 to 18 dph), and self-migration stage (19 to 270 dph).
幼鱼的顺流迁移可以分为三个阶段:漂流阶段(1至8 dph)、隐蔽阶段(9至18 dph)和自主迁移阶段(19至270 dph)。
Juveniles arrive in the estuary at 7 months, undergo osmoregulation, and grow rapidly.
幼鱼在7个月大时到达河口,进行渗透调节,并迅速生长。
At 9 months, they depart the estuary and enter the sea.
在9个月大时,它们离开河口进入大海。
Chinese paddlefish
中华匙吻鲟
The Chinese paddlefish belongs to the order Acipenseriformes, the family Polyodontidae, and the genus Psephurus in the binomial system.
中华匙吻鲟属于二名法系统中的鲟形目、匙吻鲟科和鲟属。
It was granted first-class national protected animal status in China in 1988, listed as extinct (EX) on the IUCN Red List in 2022, and listed in CITES Appendix II in 1998 (18).
它在1988年被中国列为一级国家保护动物,在2022年被国际自然保护联盟濒危物种红色名录(IUCN红色名录)列为灭绝(EX),并在1998年被列入《濒危野生动植物种国际贸易公约》附录II(18)。
The Chinese paddlefish, the longest fish in the Yangtze River, measures up to 7 m in length and weighs up to 908 kg.
中华匙吻鲟是长江中最长的鱼,体长可达7米,重量可达908公斤。

Fig. 3. Migration patterns, spawning grounds and migration paths of the five fishes. (A) Chinese sturgeon. (B) Chinese paddlefish. (C) Yangtze sturgeon. (D) Chinese sucker. (E) C. guichenoti. The five main national species, regardless of their migration patterns, body size, age of maturity, spawning patterns, and fecundity, all have a similar survival strategy of breeding in the narrow, fast, and food-poor upper reaches of the Yangtze River while feeding and growing primarily in the wide, slow, and food-rich middle and lower reaches. Their estimated extinction risk categories were based on the IUCN standards (80): CR, critically endangered; EW, extinct in the wild; EX, extinct. All photo credits: Zhenli Huang and Haiying Li.
图3. 五种鱼类的迁徙模式、产卵场和迁徙路径。(A) 中华鲟。(B) 中华匙吻鲟。(C) 长江鲟。(D) 中华鲴。(E) 鯮。这五种主要的国家物种,无论它们的迁徙模式、体型、成熟年龄、产卵模式和繁殖力如何,都有类似的生存策略:在长江狭窄、湍急和食物匮乏的上游繁殖,同时主要在宽阔、缓慢和食物丰富的中下游觅食和生长。它们估计的灭绝风险类别基于IUCN标准(80):CR,极度濒危;EW,野外灭绝;EX,灭绝。所有照片由黄镇利和李海英提供
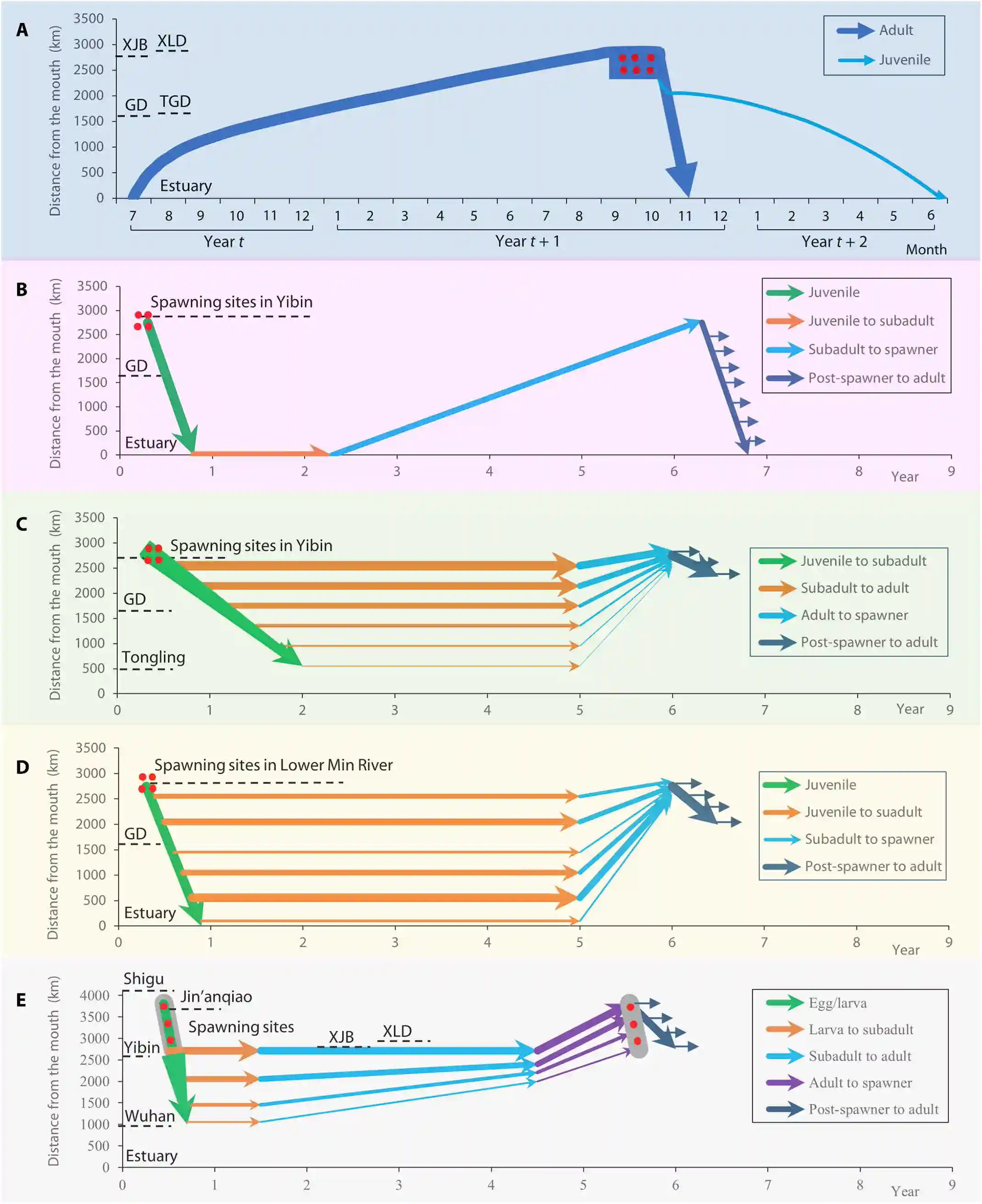
Fig. 4. Life cycles of the five fishes. (A) Chinese sturgeon (anadromy). (B) Chinese paddlefish (freshwater amphidromy). (C to E) Yangtze sturgeon, Chinese sucker, and C. guichenoti, respectively (riverine potamodromy). See text S4 for details.
图4. 五种鱼类的生命周期。(A) 中华鲟(溯河洄游性)。(B) 中华匙吻鲟(淡水两栖洄游性)。(C至E) 分别为长江鲟、中华鲴和鯮(河流性河川洄游性)。详情请参见文本S4
It mainly lives in the freshwater environment of the Yangtze River and can be found in various locations such as the mainstream, tributaries, river-connected lakes, estuary, and East China Sea (32).
它主要生活在长江的淡水环境中,可以在主流、支流、与河流相连的湖泊、河口以及东海等不同地点找到(32)。
Its spawning sites are located in the upper reaches of the Yangtze River, specifically in the lower section of the Jinsha River.
它的产卵场地位于长江上游,具体在金沙江的下游段。
In Yibin City, there are at least two spawning sites: Jiangan and Baishuxi (33).
在宜宾市,至少有两个产卵场:江安和白水溪(33)。
Baishuxi (Fig. 3B) is approximately 10 km downstream of Sankuaishi, a Chinese sturgeon spawning site. 白水溪(图3B)位于中华鲟产卵场三块石下游大约10公里处
Females lay sticky eggs on a pebbly substrate, and the breeding season occurs between March and April.
雌性在碎石基质上产下粘性的卵,繁殖季节发生在3月至4月之间。
Females typically breed between 6 and 16 years old, while males breed between 5 and 10 years old.
雌性通常在6至16岁之间繁殖,而雄性在5至10岁之间繁殖。
The sex ratio is 1:2 (34).
性别比为1:2(34)。
The existing understanding of the migration pattern and life cycle of Chinese paddlefish is erroneous, resulting in a severe underestimation of the influence of the GD on the fish (17, 18, 23, 28).
对中华匙吻鲟的迁徙模式和生命周期的现有理解是错误的,导致严重低估了葛洲坝(GD)对鱼类的影响(17, 18, 23, 28)。
To rectify this, we re-evaluated survey data to validate the migration pattern as freshwater amphidromy, and we incorporated these findings into its life cycle model (Methods and text S4.2).
为了纠正这一点,我们重新评估了调查数据,以确认其迁徙模式为淡水两栖性,并将这些发现纳入其生命周期模型(方法和文本S4.2)。
The resulting life cycle of the Chinese paddlefish is illustrated in Fig. 4B.
中华匙吻鲟的生命周期结果如图4B所示。
When males aged 5 years and females aged 6 years reach sexual maturity, they breed at Baishuxi (an example) from late March to early April.
当5岁的雄性和6岁的雌性达到性成熟时,它们在白水溪(一个例子)从3月下旬到4月上旬繁殖。
The fertilized eggs sink and adhere to pebbles and hatch into juveniles that migrate downstream and feed along nearshore waters.
受精卵下沉并附着在卵石上,孵化成幼鱼,这些幼鱼顺流而下并在近岸水域觅食。
By July and August, they reach the estuary at 4 to 5 months old.
到了7月和8月,它们在4到5个月大时到达河口。
They continue to feed and grow in estuarine and coastal areas until they become subadults.
它们继续在河口和沿海地区觅食和生长,直到成为亚成体。
The subadults reach gonadal stage II at 1.5 to 2 years old and migrate from saltwater to freshwater in the Yangtze River.
亚成体在1.5至2岁时达到性腺发育第二期,从咸水迁移到长江的淡水中。
They then begin the upriver migration to the original spawning site, occasionally entering tributaries and lakes for feeding before returning to the mainstream and moving toward the upper reaches of the river.
然后它们开始向上游的产卵场地迁移,偶尔进入支流和湖泊觅食,然后返回主流并向河流上游移动。
The migration time for subadults from the estuary to the spawning ground can range from 1 to 4 years.
亚成体从河口到产卵场的迁移时间可以从1年到4年不等。
When subadults reach gonadal stage III and are ready to breed, they migrate to the Yibin- Chongqing section during the fall and winter seasons.
当亚成体达到性腺发育第三期并准备繁殖时,它们在秋冬季节迁移到宜宾-重庆段。
After breeding, post-spawners at gonadal stage II migrate downstream and disperse throughout the habitat from Yibin to the estuary for feeding.
繁殖后,处于性腺发育第二期的产卵后个体顺流而下,并分散在整个从宜宾到河口的栖息地觅食。
Yangtze sturgeon
长江鲟
The Yangtze sturgeon belongs to the order Acipenseriformes, the family Acipenseridae, and the genus Acipenser in the binomial system.
长江鲟属于二名法系统中的鲟形目、鲟科和鲟属。
It was granted first-class national protected animal status in China in 1988, listed as extinct in the wild (EW) on the IUCN Red List in 2022, and listed in CITES Appendix II in 1998 (19).
它在1988年被中国列为一级国家保护动物,在2022年被国际自然保护联盟濒危物种红色名录(IUCN红色名录)列为野外灭绝(EW),并在1998年被列入《濒危野生动植物种国际贸易公约》附录II(19)。
The Yangtze sturgeon is mainly distributed in the upper reaches of the Yangtze River and its tributaries, including the Minjiang, Tuojiang, and Jialing rivers, with a small portion in the middle and lower reaches of the Yangtze River.
长江鲟主要分布在长江上游及其支流,包括岷江、沱江和嘉陵江,以及长江中下游的一小部分。
Spawning sites are located between the Yibin- Hejiang section, such as Anbian (Fig. 3C), Nanguang, Majiaheishibao, and Guanyintuo.
产卵场地位于宜宾-合江段之间,如岸边(图3C)、南广、马家河石堡和观音沱。
Females lay sticky eggs in spring and late fall, with most breeding occurring in spring.
雌性在春季和晚秋产下粘性的卵,大多数繁殖发生在春季。
Spawners consist of males aged 4 to 7 years and females aged 6 to 8 years, with a sex ratio of 1:2.5 (25).
产卵者由4至7岁的雄性和6至8岁的雌性组成,性别比为1:2.5(25)。
Upon conducting a thorough analysis of our proposed definition of riverine potamodromy and carefully examining the existing data, we determined that the Yangtze sturgeon can be accurately classified as a riverine potamodromous fish.
在对我们提出的河流性河川洄游性定义进行彻底分析并仔细检查现有数据后,我们确定长江鲟可以被准确地分类为河流性河川洄游性鱼类。
In addition, we collected survey data to incorporate into its life cycle model (Methods and text S4.3).
此外,我们收集了调查数据以纳入其生命周期模型(方法和文本S4.3)。
The resulting life cycle of the Yangtze sturgeon is shown in Fig. 4C.
长江鲟的生命周期结果如图4C所示。
In March- April and November- December, 4- to 7- year- old males and 6- to 8- year- old females breed in a dispersed manner along the 300- km Yibin- Hejiang section of the Yangtze River.
在3月至4月和11月至12月,4至7岁的雄性和6至8岁的雌性以分散的方式在长江宜宾-合江段300公里的范围内繁殖。
The fertilized eggs sink and adhere to gravel and hatch after 4 to 5 days.
受精卵下沉并附着在砾石上,4至5天后孵化。
At 13 dph, the larvae start feeding at the spawning sites.
在13天大时,幼体开始在产卵场地觅食。
At 30 dph, the juveniles migrate downstream along the nearshore waters to search for habitats with abundant food and to disperse and mitigate competition and predation risks.
在30天大时,幼鱼顺流而下沿着近岸水域迁移,寻找食物丰富的栖息地,并分散以减轻竞争和捕食风险。
The majority of juvenile individuals reside and forage within the mainstream and tributaries of the Luzhou- Chongqing section of the Yangtze River, while a minority migrate with the current to the middle and lower reaches of the Yangtze River, extending as far as Tongling City in Anhui Province.
大多数幼鱼个体居住并觅食在长江泸州-重庆段的主流和支流中,而少数幼鱼随水流迁移到长江中下游,远至安徽省铜陵市。
When 3- year- old males and 5- year- old females reach gonadal stage III, they begin to migrate upstream to the spawning sites in the Yibin- Hejiang section for breeding.
当3岁的雄性和5岁的雌性达到性腺发育第三期时,它们开始向上游的宜宾-合江段产卵场地迁移以进行繁殖。
During this period, which lasts at least 1 year, they occasionally enter tributaries or lakes to feed and return to the mainstream to continue their upstream journey.
在这一至少持续1年的期间,它们偶尔进入支流或湖泊觅食,然后返回主流继续它们的上游旅程。
As gonadal maturity progresses, 4- year- old males and 6- year- old females arrive at spawning sites.
随着性腺成熟度的提高,4岁的雄性和6岁的雌性到达产卵场。
After breeding, the post-spawners disperse and stay in the upper reaches for feeding.
繁殖后,产卵后的个体分散并留在上游觅食。
Once their gonads reach stage III, they migrate to the spawning sites to reproduce again.
一旦它们的性腺达到第三期,它们就迁移到产卵场再次繁殖。
Female fish usually reproduce every 2 years.
雌性鱼类通常每两年繁殖一次。
Chinese sucker
中华鲴
The Chinese sucker belongs to the order Cypriniformes, the family Catostomidae, and the genus Myxocyprinus in the binomial system.
中华鲴属于二名法系统中的鲤形目、鲴科和鲴属。
It was granted second- class national protected animal status in China in 1988 and listed as vulnerable in the China Red Data Book of Endangered Animals in 2003 (35).
它在1988年被中国列为二级国家保护动物,并在2003年被列入《中国濒危动物红皮书》中的易危物种(35)。
The Chinese sucker is mainly distributed in the Yibin- Chongqing section of the Yangtze River and the lower reaches of the Minjiang River, a major tributary of the upper Yangtze.
中华鲴主要分布在长江宜宾-重庆段和长江上游的主要支流岷江河下游。
It has also been observed in the middle and lower reaches of the Yangtze River (36).
它也在长江中下游被发现(36)。
Spawners are primarily located in the lower Minjiang River and the Yibin- Chongqing section, while juveniles and subadults are in the middle and lower Yangtze River.
产卵者主要位于岷江河下游和宜宾-重庆段,而幼体和亚成体则位于长江中下游。
Breeding occurs on gravel beaches from March to April at spawning sites such as Longxuanzi (Fig. 3D), Menkantan, Ganlongzi, Wangyemiao, Sipo, and Nanzhuatan in the lower Minjiang River.
繁殖发生在3月至4月,产卵场地位于岷江河下游的龙须滩(图3D)、门坎滩、干龙子、王家庙、四堡和南竹滩等砾石滩上。
The sex ratio is 1:1.39, and the average fecundity is 286,700 eggs.
性别比为1:1.39,平均产卵量为286,700个卵。
Historically, there were two other populations in the Yalong River and the Jialing River (37), but they were scarce in the 1970s.
历史上,雅砻江和嘉陵江还有两个其他种群,但在1970年代它们已经很稀少(37)。
The current understanding of the migration pattern and life cycle of the Chinese sucker remains uncertain (26).
目前对中华鲴的迁徙模式和生命周期的理解仍然不确定(26)。
To clarify this matter, we have undertaken a reanalysis of survey data to validate the riverine potamodromous nature of this fish’s migration.
为了澄清这个问题,我们重新分析了调查数据,以验证这种鱼的迁徙具有河流性河川洄游性。
Subsequently, we integrated the acquired data into its life cycle model (Methods and text S4.4).
随后,我们将获得的数据整合到它的生命周期模型中(方法和文本S4.4)。
The resulting life cycle of the Chinese sucker is illustrated in Fig. 4D.
中华鲴的生命周期结果如图4D所示。
Spawners (females aged 7 to 10 years; males aged 7 to 11 years) reach the Yibin section before February each year.
产卵者(7至10岁的雌性;7至11岁的雄性)每年2月前到达宜宾段。
From early February to early March, some spawners (mostly males) migrate up the Minjiang River and arrive at the Longxuanzi spawning site (an example) on the lower Minjiang River.
从2月初到3月初,一些产卵者(主要是雄性)向上游迁移到岷江,并到达岷江河下游的龙须滩产卵场(一个例子)。
In March and April, many spawners move upstream, and more females appear.
在3月和4月,许多产卵者向上游移动,更多的雌性出现。
Breeding is finished by then.
到那时,繁殖就结束了。
The fertilized eggs sink, adhere to the gravel substrate, and hatch after 10 days.
受精卵下沉,附着在砾石基质上,并在10天后孵化。
The larvae stay in the riverbed at the spawning site for 1 to 3 dph, and at 4 dph, they start feeding.
幼虫在产卵场的河床停留1至3天,然后在4天大时开始觅食。
At 19 dph, they start migrating in groups.
在19天大时,它们开始成群迁移。
Some stay in plentiful feeding areas, while others continue dispersing with the water current to suitable downstream feeding areas.
一些留在食物丰富的区域,而其他则继续随水流分散到下游合适的觅食区域。
A few reach the estuary by the current year’s fall.
一些在当年秋季到达河口。
Every spring, subadults aged 3 to 6 years with stage II gonads start their homing migration from the middle and lower reaches of the Yangtze River to the spawning sites in the lower section of the Minjiang River for at least 4 years.
每年春天,3至6岁的亚成体,具有第二期性腺的,从长江中下游开始它们的返乡迁徙到岷江河下游的产卵场,至少持续4年。
After breeding, post-spawners at gonadal stage II migrate to the mainstream and tributaries in the Yibin- Wanzhou section for feeding.
繁殖后,处于性腺发育第二期的产卵后个体迁移到宜宾-万州段的主流和支流觅食。
When their gonads reach stage III, they migrate upstream to the spawning sites to reproduce again.
当它们的性腺达到第三期时,它们向上游迁移到产卵场再次繁殖。
C. guichenoti
鯮(Coreius guichenoti)
C. guichenoti belongs to the order Cypriniformes, the family Cyprinidae, the subfamily Gobioninae, and the genus Coreius in the binomial system.
鯮属于二名法系统中的鲤形目、鲤科、鲃亚科和鯮属。
It was granted second- class national protected animal status in China in 2021.
它在2021年被中国列为二级国家保护动物。
The fish is mainly distributed in the upper and middle reaches of the Yangtze River, with a historical presence in tributaries such as the Yalong, Dadu, and Minjiang rivers (37, 38).
这种鱼主要分布在长江上游和中游,历史上也出现在雅砻江、大渡河和岷江等支流(37, 38)。
The species breeds in the rapids of the river and produces floating eggs that drift and diffuse downstream with the current.
该物种在河流的急流中繁殖,并产生随水流漂流和扩散的浮性卵。
The distribution of spawning grounds spans from Jin’anqiao in Yunnan Province to Yibin in Sichuan Province, with large spawning sites in the middle Jinsha River, such as Jin’anqiao, Duomei (Fig. 3E), and Xuzhou (Yibin).
产卵场地的分布从云南省的金安桥延伸到四川省的宜宾,金沙江中游有较大的产卵场地,如金安桥、多美(图3E)和叙州(宜宾)。
Notably, Jin’anqiao accounted for the majority, representing 84% of the total breeding size (38).
值得注意的是,金安桥占据了大多数,占总产卵规模的84%(38)。
The fish attain reproductive maturity within 3 to 7 years for males and 4 to 7 years for females, showing a minimal variance between sexes.
这种鱼在雄性3至7年、雌性4至7年内达到繁殖成熟,性别间差异很小。
The sex ratio of females to males is 1.37:1 (39, 40).
雌性与雄性的性别比为1.37:1(39, 40)。
The breeding season for this species is from late April to early July, with the peak spawning period in May to early June (40, 41).
该物种的繁殖季节从4月下旬到7月上旬,产卵高峰期在5月至6月上旬(40, 41)。
Breeding activity occurs only when the water temperature is 19° to 22°C, and the water level is rising or falling (42).
只有在水温为19°至22°C,且水位正在上升或下降时,才会发生繁殖活动(42)。
Fertilized eggs require 48 to 62 hours to hatch at water temperatures of 20.5° to 21.9°C, and the larvae start feeding 4 days after hatching (40, 43, 44).
受精卵在20.5°至21.9°C的水温下需要48至62小时孵化,幼虫在孵化后4天开始觅食(40, 43, 44)。
Previous studies on the migratory patterns of C. guichenoti have used a variety of terms and vague descriptions.
以往对鯮的迁徙模式的研究使用了各种术语和模糊的描述。
In addition, these studies have primarily focused on the eggs and/or larvae (eggs/larvae), lacking a clear description of the entire life cycle, which includes various life stages (39–44).
此外,这些研究主要关注卵和/或幼虫(卵/幼虫),缺乏对包括各个生命阶段在内的整个生命周期的清晰描述(39-44)。
To address this, we reanalyzed survey data to validate the migratory pattern as riverine potamodromy.
为了解决这个问题,我们重新分析了调查数据,以确认其迁徙模式为河流性河川洄游性。
Subsequently, we integrated these findings into the life cycle model (Methods and text S4.5).
随后,我们将这些发现整合到生命周期模型中(方法和文本S4.5)。
The resulting life cycle is shown in Fig. 4E.
产生的生命周期如图4E所示。
From April to July each year, spawners (4- to 7- year- old females and 3- to 7- year- old males) congregate at spawning sites within an approximately 1000- km stretch between Jin’anqiao and Yibin.
每年4月至7月,产卵者(4至7岁的雌性和3至7岁的雄性)聚集在金安桥到宜宾之间大约1000公里的产卵场地。
After breeding, the fertilized eggs drift with the current for approximately 1100 km to complete their embryonic development.
繁殖后,受精卵随水流漂流大约1100公里,以完成它们的胚胎发育。
This results in larvae being distributed along a 1700- km stretch between Yibin and Wuhan, which is the main feeding area for the recruitment stock (larvae, juveniles, and subadults).
这导致幼虫分布在宜宾到武汉之间1700公里的范围内,这是新招募种群(幼虫、幼鱼和亚成体)的主要觅食区。
Larvae reside and feed there, occasionally entering into neighboring tributaries or lakes for feeding.
幼虫在那里居住和觅食,偶尔进入邻近的支流或湖泊觅食。
Subadults over 1 year migrate upstream, taking at least 2 years from the Yibin- Wuhan section to the spawning sites between Jin’anqiao and Yibin.
超过1岁的亚成体向上游迁移,从宜宾-武汉段到金安桥和宜宾之间的产卵场至少需要2年时间。
They feed and grow during the migration.
它们在迁移过程中觅食和生长。
After breeding, post-spawners mainly disperse to the Jin’anqiao- Yibin section to feed and reproduce again once their gonads are mature.
繁殖后,产卵后的个体主要分散到金安桥-宜宾段去觅食,并在性腺成熟后再次繁殖。
Estimating the invalid stocks and DICs for the five fish species
估算五种鱼类的无效种群和大坝影响系数(DIC)
Dams may change the biophysical features of a river through impoundment, habitat fragmentation, blockage, alteration of the hydrological regime, hypolimnetic discharge, and nutrient retention (12, 45).
大坝可能通过蓄水、栖息地破碎、阻塞、水文情势的改变、深水层排放和营养物保持来改变河流的生物物理特征(12, 45)。
These alterations may directly influence the growth, gonadal development, migration, and reproduction of fish in the river.
这些改变可能直接影响河流中鱼类的生长、性腺发育、迁移和繁殖。
We emphasize that river closure and dam impoundment have distinct impacts on migratory fish.
我们强调河流封闭和大坝蓄水对迁徙性鱼类有独特的影响。
River closure, typically for dam construction, involves diverting water flow into a tunnel or canal, increasing flow velocity, and preventing fish from migrating upstream.
河流封闭,通常是为了建造大坝,涉及将水流分流到隧道或运河中,增加流速,并阻止鱼类向上游迁移。
Dam impoundment involves the storage of water for power generation, cutting off upstream and downstream connectivity.
大坝蓄水涉及储存水以进行发电,切断了上游和下游的连通性。
As a result, the river closure allows eggs/larvae to drift downstream but prevents upstream migration of fish below the dam.
结果,河流封闭允许卵/幼虫顺流而下,但阻止了大坝下方鱼类的上游迁移。
After impoundment, fish below the dam need facilities to move upstream, while adults above the dam cannot easily move downstream, and eggs/larvae or juveniles depend on reservoir flows and turbine structures for successful downstream passage.
蓄水后,大坝下方的鱼类需要设施来向上游移动,而大坝上方的成体不容易向下游移动,卵/幼虫或幼鱼依赖水库流动和涡轮结构来成功地向下游通过。
In the daily-regulated reservoir, such as the GD, the mean flow velocity in the reservoir exceeds the floating velocity of the eggs/larvae (0.3 m/s), allowing the majority of individuals to pass through the turbines.
在如葛洲坝这样的日调节水库中,水库中的的平均流速超过了卵/幼虫的漂浮速度(0.3米/秒),允许大多数个体通过涡轮机。
Conversely, in the seasonally regulated reservoir, such as the TGD, the flow velocity near the dam site is less than the floating velocity, resulting in the retention of all eggs/larvae within the reservoir area and subsequent mortality of underdeveloped eggs/larvae.
相反,在如三峡大坝这样的季节性调节水库中,大坝附近的流速小于漂浮速度,导致所有卵/幼虫在水库区域内滞留,随后未发育成熟的卵/幼虫死亡。
We divide the species population into spawning stock (spawners), which are sexually mature adults participating in the current year’s breeding, and recruitment stock, which includes larvae, juveniles, and subadults that have not reached the reproductive age and sexually immature adults/post-spawners that do not participate in the current year’s breeding.
我们将物种种群划分为产卵群体(产卵者),即参与当年繁殖的性成熟的成体,以及补充群体,包括未达到繁殖年龄的幼虫、幼鱼和亚成体,以及不参与当年繁殖的性未成熟的成体/产卵后个体。
Here, we define the “barrier effect” of the dam as a phenomenon where the dam cuts off upstream and downstream connectivity and changes the biophysical features of the river, preventing all or part of the spawning and recruitment stocks from completing their life cycle, especially reproduction, leading to generation of invalid stocks.
在这里,我们定义大坝的“阻碍效应”为一种现象,即大坝切断了上游和下游的连通性并改变了河流的生物物理特征,阻止了全部或部分产卵群体和补充群体完成它们的生命周期,特别是繁殖,导致产生无效种群。
We introduce the term “invalid stock” to describe the impact of dams on the migratory fish population.
我们引入了“无效种群”这个术语来描述大坝对迁徙性鱼类种群的影响。
This refers to individuals in the spawning and recruitment stocks that cannot reproduce due to the dam barrier effects.
这指的是由于大坝阻碍效应而无法繁殖的产卵群体和补充群体中的个体。
The individuals that can reproduce are referred to as “valid stock.”
能够繁殖的个体被称为“有效种群”。
Dams can render the entire or a portion of the fish population’s spawning and recruitment stocks invalid.
大坝可以使整个或部分鱼类种群的产卵群体和补充群体变为无效。
Seven distinct types of invalid stocks describe different combinations of spawning and recruitment stocks.
七种不同类型的无效种群描述了不同的产卵群体和补充群体的组合。
These types include partial spawning stock (PSS; type 1), total spawning stock (TSS, type 2), partial recruitment stock (PRS, type 3), total recruitment stock (TRS, type 4), PPS combined with PRS (PSS + PRS, type 5), PPS combined with TRS (PSS + TRS, type 6), and TSS combined with TRS (TSS + TRS, type 7) (Fig. 5A).
这些类型包括部分产卵群体(PSS;类型1)、全部产卵群体(TSS,类型2)、部分补充群体(PRS,类型3)、全部补充群体(TRS,类型4)、部分产卵群体结合部分补充群体(PSS + PRS,类型5)、部分产卵群体结合全部补充群体(PSS + TRS,类型6)和全部产卵群体结合全部补充群体(TSS + TRS,类型7)(图5A)。
One combination (TSS + PRS) cannot exist alone and instead becomes either type 2 or type 7.
一种组合(TSS + PRS)不能单独存在,而是变成类型2或类型7。
This is because if all the spawners are invalid, then there will be no reproduction and the recruitment stock will disappear.
这是因为如果所有的产卵者都无效,那么就不会发生繁殖,补充群体也会消失。
We developed a simplified model to quantify the impact of dams on data-limited migratory fish (Methods and text S3).
我们开发了一个简化模型来量化大坝对数据有限的迁徙性鱼类的影响(方法和文本S3)。
This model includes a parameter called the DIC, denoted by β, which ranges from 0 to 1.
该模型包括一个称为大坝影响系数(DIC)的参数,用β表示,范围从0到1。
A lower DIC indicates a greater barrier effect of dams on fish survival rates and the proportion of invalid stock and vice versa.
较低的DIC值表示大坝对鱼类存活率和无效种群比例的阻碍效应更大,反之亦然。
We proposed four methods to estimate the DIC: the catch method, the abundance method, the valid stock method, and the analogy method (Methods).
我们提出了四种估算DIC的方法:捕捞法、丰富度法、有效种群法和类比法(方法)。
We examined the barrier effects of the Yangtze dams on the five fishes and found that they produced all seven types of invalid stocks (movies S1 to S5).
我们检查了长江大坝对这五种鱼类的阻碍效应,发现它们产生了所有七种类型的无效种群(电影S1至S5)。
We also estimated the DICs for these fishes (text S4).
我们还估算了这些鱼类的DIC值(文本S4)。
Chinese sturgeon
中华鲟
Since 1982, this fish has not been observed breeding above the GD, and only 24.2% of the spawning stock below the GD completed breeding between 1981 and 2002 due to the mismatch between the gonad mature period (GMP) and the spawning window period (SWP) (30).
自1982年以来,这种鱼没有在葛洲坝上游观察到繁殖,1981年至2002年,由于性腺成熟期(GMP)和产卵窗口期(SWP)不匹配,葛洲坝下游只有24.2%的产卵群体完成了繁殖(30)。
As a result, 75.8% of the spawning stock was invalid (type 1) (Fig. 5B).
结果,75.8%的产卵群体是无效的(类型1)(图5B)。
After the TGD began operating in 2003, the proportion of invalid stock reached 95.5 to 100% (almost type 2) because the dam increased the water temperature by 2.7°C below the GD during the breeding season, resulting in a reduction in the SWP.
2003年三峡大坝开始运行后,由于大坝在产卵季节使葛洲坝下游的水温升高了2.7°C,导致产卵窗口期(SWP)减少,无效种群的比例达到了95.5至100%(几乎类型2)。
The operation of the Xiangjiaba and Xiluodu dams in 2013 further increased the water temperature, resulting in no overlap between the GMP and the SWP and thus entirely inhibiting breeding.
2013年,向家坝和溪洛渡大坝的运行进一步增加了水温,导致性腺成熟期(GMP)和产卵窗口期(SWP)之间没有重叠,从而完全抑制了繁殖。
Thus, the TSS became invalid (type 2).
因此,全部产卵群体变为无效(类型2)。
The cumulative impact of the cascade dams on the reproductive success of this anadromous fish has shifted the invalid stock from type 1 to type 2.
梯级大坝对这种溯河洄游性鱼类繁殖成功的累积影响已经将无效种群从类型1转变为类型2。
This indicates that effective natural breeding of the Chinese sturgeon has ceased since 2013, rendering the species functionally EX.
这表明自2013年以来,中华鲟的有效自然繁殖已经停止,使得该物种在功能上灭绝(EX)。
The “barrier effects” of the Yangtze dams on the Chinese sturgeon are manifested in delayed gonadal development by shortening the migration distance and inhibiting reproduction by elevated water temperatures (30).
长江大坝对中华鲟的“阻碍效应”表现在通过缩短迁移距离延迟性腺发育,并通过提高水温抑制繁殖(30)。
We used the abundance method to estimate the Chinese sturgeon DICs corresponding to the cumulative impact
我们使用丰富度法估算了对应于累积影响的中华鲟的大坝影响系数(DIC)
of the cascade dams during different periods. This method relied on annual population size data (30). In the spawning stock, females were 13 to 34 years old and males were 8 to 27 years old. Therefore, the impact of the GD on males and females began in 1989 and 1994, respectively, while the impact of the TGD started after its impoundment in June 2003 and that of the Xiluodu Dam began in 2013.
在不同时期,梯级大坝的累积影响。这种方法依赖于年度种群大小数据(30)。在产卵群体中,雌性的年龄在13至34岁之间,雄性在8至27岁之间。因此,葛洲坝(GD)对雄性和雌性的影响分别从1989年和1994年开始,而三峡大坝(TGD)的影响始于2003年6月蓄水后,溪洛渡大坝的影响则从2013年开始。
Table 1 shows the DICs for the spawners corresponding to the GD, TGD, and Xiluodu Dam. The β value of GD is 0.85 for males and 0.92 for females, indicating that the GD has more impact on males than females. The increasing effect of the Yangtze dams on the spawners, corresponding to β = 0.88 to 0.93, indicates that the Chinese sturgeon has high life history plasticity, with a recession rate of approximately 10% per year.
表1显示了对应于葛洲坝(GD)、三峡大坝(TGD)和溪洛渡大坝的产卵者的DIC值。GD的β值对雄性为0.85,对雌性为0.92,表明GD对雄性的影响大于雌性。长江大坝对产卵者的影响逐渐增加,对应的β值为0.88至0.93,表明中华鲟具有高度的生活史可塑性,每年衰退率约为10%。
Before 1981, the marine stock was above 30,000 individuals. Only approximately 5% of the marine stock enters the Yangtze River each year (30). As a result, the population has been declining for more than 40 years.
在1981年之前,海洋种群数量超过30,000个个体。每年大约有5%的海洋种群进入长江(30)。因此,种群数量已经下降了40多年。
Chinese paddlefish
中华匙吻鲟
Before the river closure of the GD in January 1981, Chinese paddlefish spawners and partial subadults migrated to the upper Yangtze River during the fall and winter of 1980 (Fig. 5C).
在1981年1月葛洲坝(GD)的河流封闭之前,中华匙吻鲟的产卵者和部分亚成体在1980年的秋季和冬季迁移到了长江上游(图5C)。
The spawners, known as the “old” spawning stock, were 6 to 16 years old for females and 5 to 10 years old for males.
产卵者,被称为“旧”产卵群体,雌性的年龄在6至16岁之间,雄性在5至10岁之间
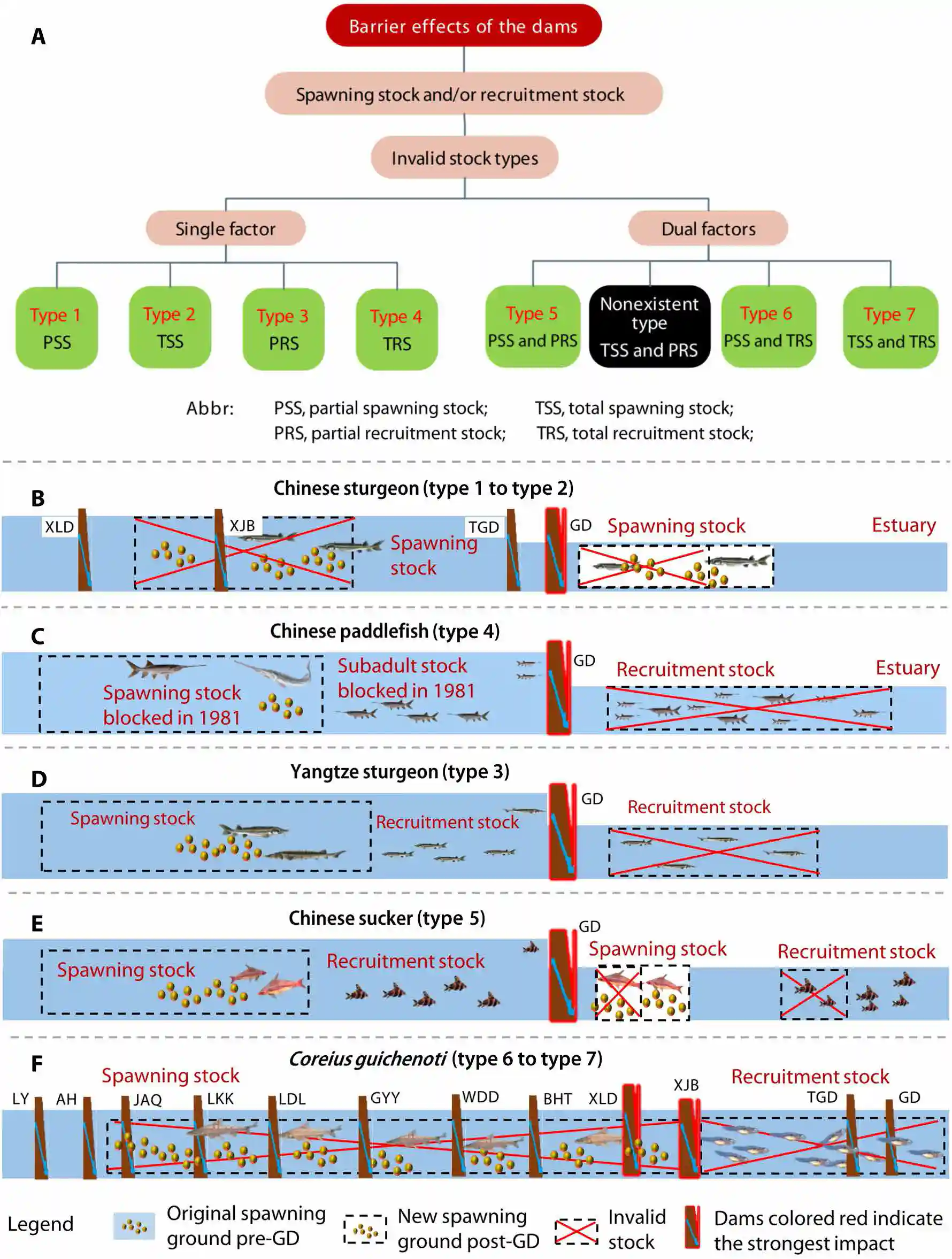
Fig. 5. Seven types of invalid stocks created by the barrier effects of the 12 Yangtze dams on the five fishes. (A) Seven types of invalid stocks. (B to F) Schematic showing the barrier effects of dams on the five fishes. In the panel: GD; TGD; XJB; XLd; BHt, Baihetan dam; Wdd, Wudongde dam; GYY, Guanyinyan dam; LdL, Ludila dam; LKK, Longkaikou dam; JAQ, Jin’anqiao dam; AH, Ahai dam; LY, Liyuan dam.
图5. 12座长江大坝的阻碍效应在五种鱼类上创造的七种无效种群类型。(A) 七种无效种群类型。(B至F) 示意图,展示了大坝对五种鱼类的阻碍效应。在图中:GD;TGD;XJB;XLd;BHt,白鹤滩大坝;Wdd,乌东德大坝;GYY,观音岩大坝;LdL,鲁地拉大坝;LKK,龙开口大坝;JAQ,金安桥大坝;AH,阿海大坝;LY,梨园大坝
Table 1. DICs for the five fishes in the Yangtze River. In the table, GD, Gezhouba Dam; TGD, Three Gorges Dam; XJB/XLD, Xiangjiaba Dam/Xiluodu Dam.Parentheses indicate the applicable DIC period. See text S4 for details. N/A, not applicable.
表1:长江中五种鱼类的DIC。表中,GD,葛洲坝;三峡大坝;XJB/XLD,向家坝/溪洛渡大坝。括号表示适用的DIC周期。详见正文S4。N/A,不适用
| Fish species鱼类 | Dam水坝 | GD | TGP | XJB/XLD |
|---|---|---|---|---|
| Chinese sturgeon中华鲟 | 0.93 (1989-2002) for spawners产卵者; 0.882 (2003-2012) for spawners; 0.881 (2013-) for spawners; 0.842 | 0.85 (1989-2002) for males男性; 0.92 | 0.976 (2003-2012) for males; 0.842 (2013-) for males; 0.911(2013-) for females | |
| Chinese paddlefish白鲟 | 0.63 (1981-1984) for the old spawning stock; 0.78 (1985-) for the new spawning stock新产卵种群 | N/A | N/A | |
| Yangtze sturgeon长江鲟 | 0.75 (1981-) | N/A | N/A | |
| Chinese sucker胭脂鱼 | 0.85 for spawners above the GD (1981-2009) and below the GD | 0.98 for spawners (2010-) and juveniles少年 (2003-) above the TGD; 0.74 (1985-2002), for juveniles (1981-2002) | N/A | |
| Coreius guichenoti圆口铜鱼 | 0.83 for eggs/larvae幼体(1981-2002) above the GD; 0.8 for subadults and adults below the GD (1981-2002) | 0.98 for eggs/larvae (2003-2007) above the TGD; 0.32 for subadults and larvae above the TGD (2008-) | 0.53 (2008-) for spawners and eggs/larvae above the TGD |
Partial subadults, 2 to 5 years old, known as the “new” spawning stock, represented a small proportion of the subadults.
部分2至5岁的亚成体,被称为“新”产卵群体,代表了亚成体中的一小部分。
The majority of the subadults were blocked by the GD’s river closure and remained below the dam.
大多数亚成体被GD的河流封闭所阻挡,留在了大坝下游。
Therefore, breeding activities of the Chinese paddlefish in the Yangtze River were conducted by the old stock until 1985, when they were gradually replaced by the new stock.
因此,长江中华匙吻鲟的繁殖活动由旧群体进行,直到1985年逐渐被新群体取代。
After 1981, all juveniles that hatched in the upper Yangtze had to migrate downstream through the turbines of the GD.
1981年之后,所有在长江上游孵化的幼鱼必须通过葛洲坝的涡轮机向下游迁移。
As a result, the TRS was invalid (type 4) because the GD prevented upriver migration, meaning successful breeding above the GD did not supplement the spawning stock.
结果,全部补充群体(TRS)变为无效(类型4),因为GD阻止了上游迁移,这意味着在GD上游的成功繁殖并没有补充产卵群体。
The barrier effect of the GD on the Chinese paddlefish is evidenced by the need for newly hatched juveniles to pass through the dam’s turbines and the obstruction of their return to their natal habitat.
GD对中华匙吻鲟的阻碍效应表现在新孵化的幼鱼需要通过大坝的涡轮机,以及它们返回原生栖息地的障碍。
In addition, the inability of the fish to reproduce below the dam exacerbates the adverse effects.
此外,鱼类无法在大坝下游繁殖加剧了不利影响。
Thus, the Chinese paddlefish has sealed its fate of inevitable extinction from a life cycle perspective after 1981, relying solely on the new and old spawning stocks above the dam to survive for another decade.
因此,从生命周期的角度来看,中华匙吻鲟在1981年后注定要灭绝,只能依靠大坝上游的新旧产卵群体再生存十年。
We used the catch method to estimate the DICs of Chinese paddlefish. Wei et al. (46) documented the quantity of fish captured below the GD between 1981 and 1995.
我们使用捕捞法估算中华匙吻鲟的DIC值。魏等人(46)记录了1981年至1995年间在GD下游捕获的鱼量。
The captured fish had successfully navigated downstream through hydraulic turbines from their upstream spawning grounds, thus providing insight into fluctuations in the spawning stock size.
被捕捞的鱼成功地从上游产卵地产卵后通过液压涡轮机向下游迁移,从而为了解产卵群体大小的波动提供了见解。
As shown in fig. S2, the pre-1985 population decline was primarily due to the declining old stock, whereas the post-1985 decline was driven by the new stock.
如图S2所示,1985年前的种群下降主要是由于旧群体的减少,而1985年后的下降则是由新群体驱动的。
Using the simplified model, we performed a regression analysis on the pre- and post-1985 data, yielding the DICs of β = 0.63 (1981–1984) and β = 0.78 (1985–) (Table 1).
使用简化模型,我们对1985年前和1985年后的数据进行了回归分析,得出了β值为0.63(1981-1984年)和β值为0.78(1985年以后)(表1)的DIC。
The Chinese paddlefish had a DIC of 0.63 to 0.78, while the Chinese sturgeon had a DIC of 0.88 to 0.93, indicating that the paddlefish is more affected by the GD and has lower life history plasticity.
中华匙吻鲟的DIC值为0.63至0.78,而中华鲟的DIC值为0.88至0.93,表明匙吻鲟受GD的影响更大,且生活史可塑性较低。
Yangtze sturgeon
长江鲟
We assumed a linear distribution of population density along the migration route, reflecting a decrease in newly hatched juveniles as they moved downstream.
我们假设沿迁徙路线的种群密度呈线性分布,反映了新孵化的幼鱼在向下游移动时数量的减少。
Approximately 75% of the annual juveniles were distributed upstream of the GD and 25% downstream.
大约75%的年度幼鱼分布在葛洲坝(GD)上游,25%分布在下游。
Figure 6A shows the density distribution of juveniles pre- and post-GD.
图6A显示了葛洲坝建成前后幼鱼的密度分布。
The dotted lines at the GD site indicate that approximately 30% of the juveniles passing through the turbines were killed, while others survived and reached the middle and lower reaches of the Yangtze River.
葛洲坝现场的虚线表明,大约30%通过涡轮机的幼鱼被杀死,而其他存活下来并到达了长江的中下游。
Since 1981, an estimated 25% of the juveniles that hatched in the upstream spawning grounds annually have been unable to migrate back upstream for reproduction or successfully reproduce below the dam.
自1981年以来,估计每年在上游产卵地产卵孵化的幼鱼中有25%无法返回上游进行繁殖或在大坝下游成功繁殖。
This has resulted in them becoming an invalid stock (type 3) (Fig. 5D).
这导致它们成为了无效种群(类型3)(图5D)。
The barrier effect of the GD has caused the invalidation of 25% of the recruitment stock each year since 1981, leading to a persistent decline in population.
葛洲坝的阻碍效应自1981年以来每年导致25%的补充群体失效,导致种群数量持续下降。
Using the valid stock method, we estimated the DIC of juveniles to be β = 0.75, as shown in Fig. 6A.
使用有效种群法,我们估算了幼鱼的DIC值为β=0.75,如图6A所示。
Under the assumption that the GD has an equivalent effect on both the spawning stock and the recruitment stock, we can derive the DIC of the entire population of Yangtze sturgeon (Table 1).
在假设葛洲坝对产卵群体和补充群体具有相同影响的前提下,我们可以推导出长江鲟整个种群的DIC值(表1)。
Chinese sucker
中华鲴
Figure 6B (solid red line) shows that juveniles were mainly distributed along a 500-km stretch between Yibin and Chongqing and a 1675-km stretch between Yichang and the estuary.
图6B(实红线)显示,幼鱼主要分布在宜宾和重庆之间的500公里范围内,以及宜昌和河口之间的1675公里范围内。
The Three Gorges section was just a migration corridor.
三峡段仅仅是一个迁徙通道。
Assuming an even distribution, we estimated that the number of juveniles in the middle and lower reaches of the Yangtze River was roughly three times the number in the upper reaches (75:25%).
假设分布均匀,我们估计长江中下游的幼鱼数量大约是上游数量的三倍(75:25%)。
As 3- to 6- year- old subadults (solid blue line) moved closer to upstream spawning sites, their density increased.
当3至6岁的亚成体(实蓝线)更接近上游产卵地时,它们的密度增加。
Approximately 65% of subadults were found in the upper reaches, whereas 35% were in the middle and lower reaches.
大约65%的亚成体发现在上游,而35%在中下游。
Spawners were mainly located in the 500-km river section between Yibin and Chongqing, with a higher density in Yibin and a lower density in Chongqing (solid black line).
产卵者主要位于宜宾和重庆之间的500公里河段,宜宾的密度较高,重庆的密度较低(实黑线)。
Since the river closure for the GD on 4 January 1981, the Chinese sucker population has split into two independent subpopulations.
自1981年1月4日葛洲坝(GD)的河流封闭以来,中华鲴的种群已经分裂成两个独立的亚种群。
The upstream subpopulation continued to reproduce but lost subadults below the GD (blue dotted line), causing a decrease in breeding size (black dotted line on the left side).
上游亚种群继续繁殖,但失去了GD下游的亚成体(蓝色虚线),导致繁殖规模减少(左侧黑色虚线)。
In addition, approximately 30% of the juveniles passing through the turbines were killed each year, resulting in a decrease in juvenile densities above and below the GD.
此外,每年大约有30%通过涡轮机的幼鱼被杀死,导致GD上游和下游的幼鱼密度减少。
The 3- year- old subadults below the GD would reach reproductive age in 1985.
GD下游的3岁亚成体将在1985年达到繁殖年龄。
Only 74% of the spawners below the GD reproduced, and 26% became invalid.
GD下游只有74%的产卵者进行了繁殖,而26%变得无效。
The barrier effects of the GD on the Chinese sucker are manifested in the loss of juveniles passing through the turbines each year and in the invalid stock of partial spawners below the dam (Fig. 5E).
GD对中华鲴的阻碍效应表现在每年通过涡轮机的幼鱼的损失以及大坝下游部分产卵者无效种群的损失(图5E)。
The combination of these two factors was classified as type 5.
这两个因素的结合被归类为类型5。
Jiang and Yu (47) reported 153 bycatch individuals in the Tongling section of Anhui Province from 1997 to 2002.
江和于(47)报告了1997年至2002年在安徽省铜陵段捕获的153个兼捕个体。
Using the catch method, we estimated a DIC of 0.85 for spawners (1985–2002) and juveniles (1981–2002) below the GD (fig. S3).
使用捕捞法,我们估算了GD下游产卵者(1985-2002年)和幼鱼(1981-2002年)的DIC值为0.85(图S3)。
Before 1981 (pre-GD), Chinese suckers bred only in the upper reaches, and there was no evidence of breeding in the middle and lower reaches.
在1981年之前(GD前),中华鲴只在上游繁殖,没有中下游繁殖的证据。
After 1981 (post-GD), Chinese suckers were forced to breed below the GD, but the unfavorable environmental conditions prevented some spawners from reproducing successfully.
1981年之后(GD后),中华鲴被迫在GD下游繁殖,但不利的环境条件阻止了一些产卵者成功繁殖。
We estimated that the breeding efficiency of Chinese suckers below the GD was 0.74, meaning that only 74% of the spawners participated in breeding activities each year.
我们估计GD下游中华鲴的繁殖效率为0.74,这意味着每年只有74%的产卵者参与繁殖活动。
After 2003 (post-TGD), no upstream juveniles were received below the GD.
2003年之后(TGD后),下游没有收到上游的幼鱼。
The DICs below the GD decreased from 0.85 for spawners (1985–2002) and juveniles (1981–2002) to 0.74 for spawners and juveniles (2003–).
GD下游的DIC值从产卵者(1985-2002年)和幼鱼(1981-2002年)的0.85下降到产卵者和幼鱼(2003年以后)的0.74。
There was a lack of relevant data for the upper reaches of the Yangtze River.
长江上游缺乏相关数据。
We then assumed that the DIC for spawners above the GD/TGD was the same as below the GD, with a value of 0.85.
然后我们假设GD/TGD上游的产卵者的DIC与下游相同,值为0.85。
From 2003 onward, newly hatched juveniles remained above the TGD and started to reproduce in 2009.
从2003年起,新孵化的幼鱼留在了TGD上游,并在2009年开始繁殖。
To account for the potential mortality of unsuitable juveniles in the lentic water environment of the reservoir, we adjusted the DIC to 0.98.
为了考虑水库静水环境中不适宜幼鱼的潜在死亡率,我们将DIC调整为0.98。
Consequently, DICs above the GD/TGD for spawners were 0.85 (1981–2009) and 0.98 (2010–), while DICs for juveniles were 0.85 (1981–2002) and 0.98 (2003–) (Table 1).
因此,GD/TGD上游产卵者的DIC值分别为0.85(1981-2009年)和0.98(2010年以后),而幼鱼的DIC值分别为0.85(1981-2002年)和0.98(2003年以后)(表1)。
C. guichenoti
鯮(Coreius guichenoti)
In Fig. 6C, we assume a linear density distribution of spawners from Jin’anqiao to Yibin.
在图6C中,我们假设从金安桥到宜宾的产卵者的密度呈线性分布
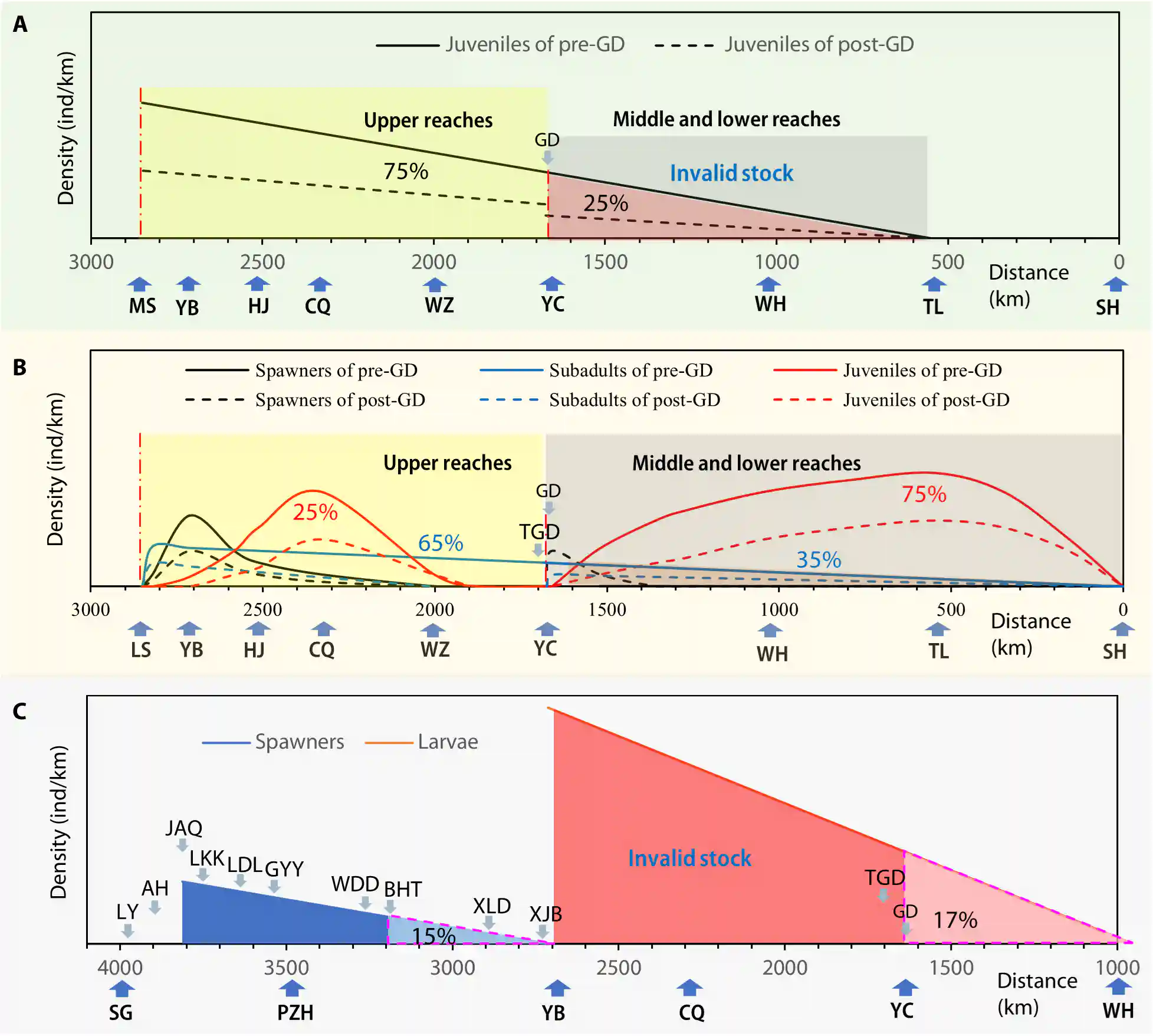
Fig. 6. Schematic showing the population density distributions of three riverine potamodromous species in the Yangtze River before and after the construction of the Yangtze dams. (A) Yangtze sturgeon. (B) Chinese sucker. (C) Coreius guichenoti. The distance upriver from the mouth of the Yangtze River is plotted on the abscissa, with geographically iconic places indicated by deep blue arrows. In Yunnan Province: SG, Shigu town. In Sichuan Province: PZH, Panzhihua city; LS, Leshan city; MS, Maoshui town; YB, Yibin city; HJ, Hejiang county. In Chongqing Municipality: CQ, downtown Chongqing; WZ, Wanzhou district. In Hubei Province: YC, Yichang city; WH, Wuhan city. In Anhui Province: TL, Tongling city. In Shanghai Municipality: SH, Shanghai city. In the (A) to (C), the dam locations are indicated by light blue arrows: GD, TGD, XJB, XLd, BHt, Wdd, GYY, LdL, LKK, JAQ, AH, and LY. See text S4 for details.
图6. 示意图,展示了长江大坝建设前后长江中三种河流性河川洄游性物种的种群密度分布。(A) 长江鲟。(B) 中华鲴。(C) 鯮。从长江口起向上游的距离在横轴上表示,地理上标志性的地方由深蓝色箭头指示。在云南省:SG,石鼓镇。在四川省:PZH,攀枝花市;LS,乐山市;MS,茅石镇;YB,宜宾市;HJ,合江县。在重庆市:CQ,重庆市区;WZ,万州区。在湖北省:YC,宜昌市;WH,武汉市。在安徽省:TL,铜陵市。在上海市:SH,上海市。在(A)至(C)中,大坝的位置由浅蓝色箭头指示:GD,TGD,XJB,XLd,BHt,Wdd,GYY,LdL,LKK,JAQ,AH和LY。详情请参见文本S4
The Jin’anqiao spawning site was the largest, with the others decreasing linearly in size.
金安桥产卵场是最大的,其他产卵场的大小线性递减。
Consequently, the Yibin section has the highest larval density, correlating with a migration distance of approximately 1100 km, during which fertilized eggs undergo development into feeding larvae and remain in the section.
因此,宜宾段的幼虫密度最高,这与大约1100公里的迁移距离相关,在这个距离中,受精卵发育成觅食的幼虫并留在该段。
Approximately 17% of the eggs/larvae below the GD and TGD were considered invalid stock (type 3), suggesting that these two dams had less impact on the species (Fig. 5F).
GD和TGD下游大约17%的卵/幼虫被视为无效种群(类型3),表明这两个大坝对物种的影响较小(图5F)。
However, the Xiangjiaba and Xiluodu dams, located exactly on the demarcation line between the spawning ground and the nursery habitat, completely separated the spawning stock and the recruitment stock, disrupting their life cycle integrity.
然而,向家坝和溪洛渡大坝正好位于产卵场和育幼场之间的分界线上,完全隔离了产卵群体和补充群体,破坏了它们的生命周期完整性。
As a result, the minority of the spawning stock and the TRS became invalid (type 6).
结果,少数的产卵群体和全部补充群体(TRS)变得无效(类型6)。
In addition, following the operation of the 12 major dams, the TSS and the TRS also became invalid (type 7).
此外,在12座主要大坝运行后,全部产卵群体(TSS)和全部补充群体(TRS)也变得无效(类型7)。
The population collapse of C. guichenoti was not due to the barrier effect of a single dam, but rather the superposition of multiple barrier effects from the Yangtze dams, manifested in increasing proportions of invalid stocks in both the spawning and recruitment stocks as the number of cascade dams increased.
鯮(Coreius guichenoti)的种群崩溃不是由单个大坝的阻碍效应引起的,而是由于长江大坝的多重阻碍效应的叠加,随着梯级大坝数量的增加,产卵群体和补充群体中无效种群的比例不断增加。
We used the four methods to estimate the DICs for C. guichenoti to understand how a combination of methods can be used under different data scenarios as follows (Table 1):
我们使用了四种方法来估算鯮(Coreius guichenoti)的DIC值,以了解在不同数据情景下如何使用这些方法的组合(表1):
In valid stock method, Fig. 6C shows that 17% of the total eggs/ larvae were below the GD.
有效种群法中,图6C显示GD下游有17%的卵/幼虫总数。
After 1981 (post-GD), this 17% became invalid stock because they were unable to reproduce below the GD as they matured.
1981年之后(GD后),这17%变成了无效种群,因为它们在成熟后无法在GD下游繁殖。
The remaining 83% above the GD were considered valid stock.
GD上游剩余的83%被认为是有效种群。
Therefore, the DIC of the eggs/larvae above the GD was 0.83 (1981–2002).
因此,GD上游卵/幼虫的DIC值为0.83(1981-2002年)。
After 2003, the TGD prevented eggs/larvae from passing through, causing an increase in their density in the reservoir.
2003年之后,TGD阻止了卵/幼虫通过,导致它们在水库中的密度增加。
Taking into account minor mortality in the reservoir, the DIC of eggs/ larvae above the TGD was estimated to be 0.98 (2003–2007).
考虑到水库中的轻微死亡,TGD上游卵/幼虫的DIC值估计为0.98(2003-2007年)。
In abundance method, since 1981, a survey of egg-eating fish below the GD has been used to study the reproductive behavior of Chinese sturgeon, as C. guichenoti is a major predator of their eggs.
丰富度法中,自1981年以来,对GD下游的食卵鱼进行调查,以研究中华鲟的繁殖行为,因为鯮是其卵的主要捕食者。
Yu et al. (48) provided data on the estimated number of subadults/adults of C. guichenoti from 1997 to 2001, allowing us to calculate a DIC of 0.8 (1981–2002) (fig. S4A).
于等人(48)提供了1997年至2001年鯮(Coreius guichenoti)亚成体/成体的估计数量数据,使我们能够计算出0.8的DIC值(1981-2002年)(图S4A)。
In catch method, Tao et al. (49) reported catch numbers below the GD from 2005 to 2007, indicating a changing trend in the C. guichenoti stocks (subadults/adults) after 2003 (post-TGD), with a DIC of 0.32 (2003–) (fig. S4B).
在捕捞法中,陶等人(49)报告了2005年至2007年GD下游的捕捞数量,表明2003年之后(TGD后)鯮(Coreius guichenoti)种群(亚成体/成体)的趋势发生了变化,DIC值为0.32(2003年以后)(图S4B)。
In 2007–2008, the XJB and XLD’s river closures resulted in the disappearance of spawning grounds in the two reservoirs, and 15% of the spawners became invalid stock (Fig. 6C).
2007-2008年,向家坝(XJB)和溪洛渡大坝(XLD)的河流封闭导致两个水库的产卵场消失,15%的产卵者变成了无效种群(图6C)。
We consider the Xiangjiaba and Xiluodu dams as one unit due to their proximity and the 1- year difference in their completion dates.
由于向家坝和溪洛渡大坝的位置接近以及它们完工日期相差一年,我们将它们视为一个单位。
Here, we propose two approaches to estimate the DICs for C. guichenoti affected by these dams.
在这里,我们提出了两种估算受这些大坝影响的鯮(Coreius guichenoti)的DIC值的方法。
In analogy method, the Chinese paddlefish spawners above the GD were totally valid (β = 0.63), and 85% of the spawners above the Xiangjiaba and Xiluodu dams were valid (Fig. 6C).
在类比法中,GD上游的中华匙吻鲟产卵者完全有效(β=0.63),向家坝和溪洛渡大坝上游85%的产卵者有效(图6C)。
Using this information, we estimated the DIC of spawners and eggs/larvae to be 0.54 (0.63 × 0.85), as they are equivalent in this context.
利用这些信息,我们估算了产卵者和卵/幼虫的DIC值为0.54(0.63×0.85),因为在这种情况下它们是等效的。
In catch method, Tang et al. (38) analyzed the survey data of eggs/ larvae in the Geliping section of Panzhihua from 2006 to 2010, showing the effect of Xiangjiaba and Xiluodu dams on spawners.
在捕捞法中,唐等人(38)分析了2006年至2010年攀枝花格里坪段的卵/幼虫调查数据,显示了向家坝和溪洛渡大坝对产卵者的影响。
With a simplified model, the regression analysis yielded a DIC of 0.53 for spawning stock and eggs/larvae (fig. S4C).
使用简化模型,回归分析得出产卵群体和卵/幼虫的DIC值为0.53(图S4C)。
The results indicate that the DIC is 0.54 for the analogy method and 0.53 for the catch method, both of which are reasonable.
结果表明,类比法的DIC值为0.54,捕捞法的DIC值为0.53,这两个值都是合理的。
Thus, we chose the DIC value of 0.53 (2008–).
因此,我们选择了0.53的DIC值(2008年以后)。
Trend analysis of dam-induced impacts using invalid stock types
使用无效种群类型对大坝引起的影响进行趋势分析
We investigated the correlation between the invalid stock and the DIC.
我们研究了无效种群和DIC之间的相关性。
Our results suggest that invalid stock types can indicate the trend of dam-induced effects.
我们的结果显示,无效种群类型可以指示大坝引起的效应趋势。
Identifying these types helps to estimate the magnitude of impacts and compare them with data-limited fish.
识别这些类型有助于估计影响的大小并与数据有限的鱼类进行比较。
Population decline accelerates as the proportion of invalid stock increases.
随着无效种群比例的增加,种群下降加速。
Figure 7A shows that total invalidation (type 2, type 4, and type 7) causes larger population declines than partial invalidation (type 1 and type 3).
图7A显示,完全无效(类型2、类型4和类型7)导致的种群下降大于部分无效(类型1和类型3)。
Despite their different species and migration patterns, both Yangtze sturgeon and Chinese paddlefish show a consistent trend where type 3 (PRS) has a higher DIC of 0.75 compared to type 4 (TRS) with a value of 0.63.
尽管物种和迁徙模式不同,长江鲟和中华匙吻鲟都显示出一致的趋势,其中类型3(部分补充群体PRS)的DIC值较高,为0.75,而类型4(全部补充群体TRS)的值为0.63。
Protecting recruitment stock is more important than protecting spawning stock.
保护补充群体比保护产卵群体更重要。
Figure 7B shows that the DICs for recruitment stock cases (type 3 and type 4) were lower (0.75 and 0.63) than for spawning stock cases (type 1 and type 2) (0.93 and 0.882).
图7B显示,补充群体案例(类型3和类型4)的DIC值(0.75和0.63)低于产卵群体案例(类型1和类型2)(0.93和0.882)。
This trend was also observed in dual-factor cases.
这一趋势也在双因素案例中观察到。
This finding is unexpected as the prevailing viewpoint prioritizes protecting spawning stock.
这一发现出乎意料,因为主流观点优先保护产卵群体。
The cascade dams accelerate the rate of population decline to extinction.
梯级大坝加速了种群下降至灭绝的速率。
Figure 7C shows that the Chinese sturgeon population declined at an accelerated rate, as the DICs of GD, TGD, and other upper cascade dams were 0.93, 0.882, and 0.881, respectively.
图7C显示,中华鲟的种群以加速的速率下降,因为葛洲坝(GD)、三峡大坝(TGD)和其他上游梯级大坝的DIC值分别为0.93、0.882和0.881。
For C. guichenoti, the TGD had a slight positive effect.
对于鯮(Coreius guichenoti),三峡大坝(TGD)有轻微的积极效应。
However, when other upper dams, such as the Xiangjia and Xiluodu dams were built, the fish population declined by almost half each year.
然而,当建造了其他上游大坝,如向家坝和溪洛渡大坝时,鱼类种群数量几乎每年减少一半。
In addition, different dams exerted distinct effects on the population of C. guichenoti.
此外,不同的大坝对鯮(Coreius guichenoti)的种群产生了不同的影响。
The Xiangjiaba and Xiluodu dams have the greatest impact (β = 0.53), followed by the GD (0.88) and TGD (0.98).
向家坝和溪洛渡大坝的影响最大(β=0.53),其次是葛洲坝(0.88)和三峡大坝(0.98)。
Notably, the combination of TSS and TRS (type 7) represents the worst-case scenario for this species, as shown in Fig. 7D, with the lowest DIC (0.32) indicating the fastest population decline.
值得注意的是,全部产卵群体(TSS)和全部补充群体(TRS)的结合(类型7)代表了这一物种的最坏情况,如图7D所示,最低的DIC值(0.32)表明种群下降速度最快。
On the basis of the data from five species and 12 Yangtze dams, we conclude that further research on the invalid stock concept will provide a larger database to evaluate the impacts of dams on data-limited fish.
基于五种物种和12座长江大坝的数据,我们得出结论,对无效种群概念的进一步研究将提供一个更大的数据库,以评估大坝对数据有限的鱼类的影响。
We can extend the invalid stock concept to analyze the long-term impact of environmental factors such as climate change and water pollution on migratory fish.
我们可以扩展无效种群概念,以分析气候变化和水污染等环境因素对迁徙性鱼类的长期影响。
By conducting field and indoor experiments, we can assess the effects of high water temperatures or pollution on reproduction.
通过进行野外和室内实验,我们可以评估高温或污染对繁殖的影响。
Recreating population decline processes of the five fish species
重现五种鱼类种群下降过程
We have completed the validation of the simplified model based mainly on the Chinese sturgeon case (Methods).
我们已经基于中华鲟案例(方法)完成了简化模型的验证。
To use this model, we need to determine the initial population sizes of the species before the dam was built.
要使用这个模型,我们需要确定建坝前物种的初始种群大小。
The estimation methods depend on the availability of specific data, as data limitations vary.
估算方法取决于特定数据的可用性,因为数据限制各不相同。
Now, we have very limited information on the abundance of the four fish species besides the Chinese sturgeon.
现在,除了中华鲟之外,我们对其他四种鱼类的丰富度信息非常有限。
There are two main categories of data available for analysis.
可供分析的数据主要分为两大类。
The first is catch statistics, which includes Chinese paddlefish, Yangtze sturgeon, and Chinese sucker.
第一类是捕捞统计数据,包括中华匙吻鲟、长江鲟和中华鲴。
Using an analogy method, we assumed that the exploitation rate of these three species mirrored that of Chinese sturgeon.
使用类比法,我们假设这三种物种的利用率与中华鲟相似。
Consequently, we used the amount of catch to estimate the population size.
因此,我们使用捕捞量来估计种群大小。
The second category includes statistics on eggs/larvae for C. guichenoti.
第二类包括鯮(Coreius guichenoti)的卵/幼虫统计数据
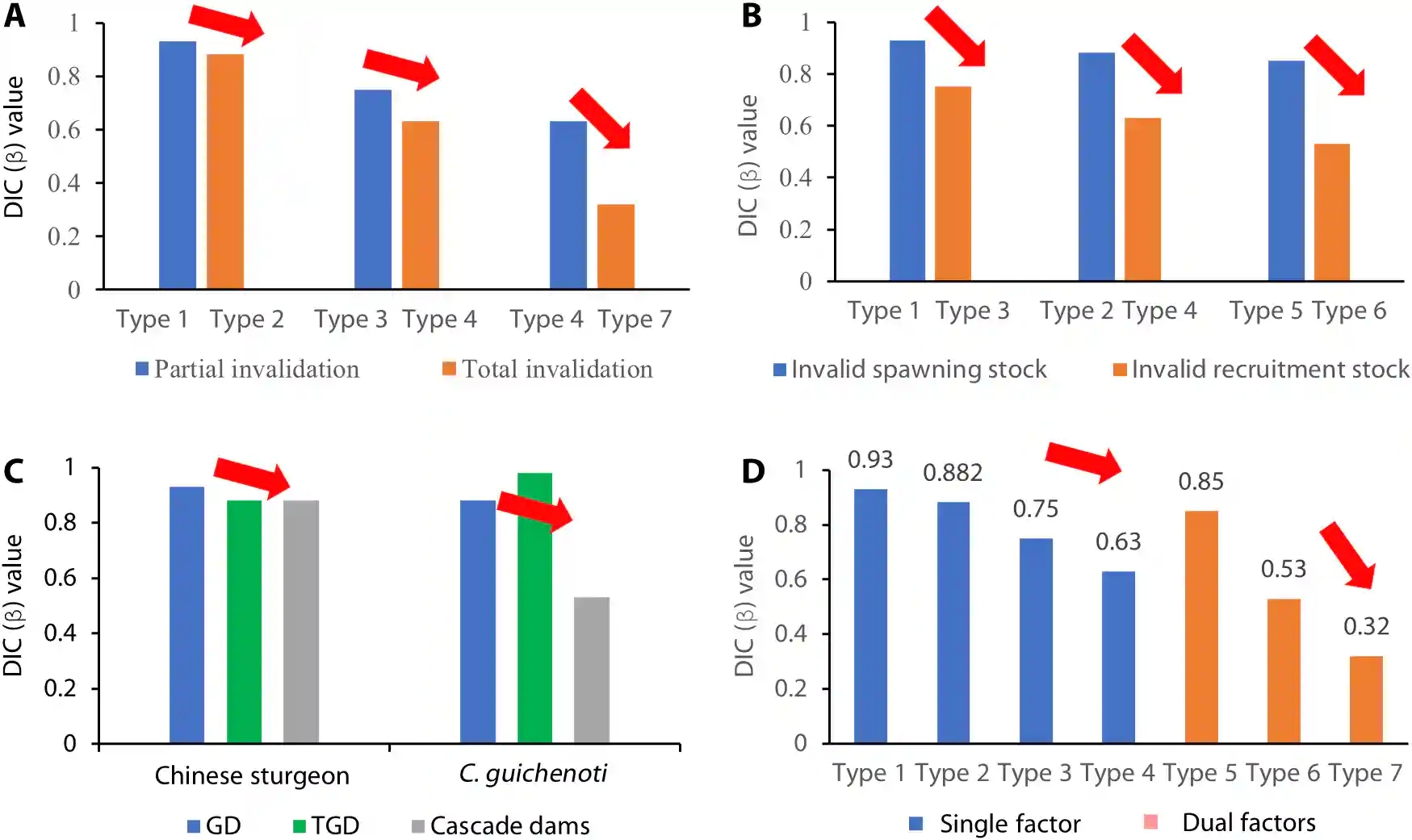
Fig. 7. Invalid stock types reflecting the trends of dam-induced impacts. (A) Comparison of DICs for partially invalid stocks and totally invalid stocks. (B) Comparison of the DICs for invalid spawning and recruitment stocks. (C) Comparison of the DICs for the GD, TGD, and the other cascade dams. (D) DICs of various invalid stock types.
图7. 反映大坝引起影响趋势的无效种群类型。(A) 部分无效种群和完全无效种群的DIC比较。(B) 无效产卵种群和补充种群的DIC比较。(C) GD、TGD和其他梯级大坝的DIC比较。(D) 各种无效种群类型的DIC
Given the number of eggs/larvae at a particular station in a river, we extrapolated the total for that year using the eggs/larvae density distribution diagram along the river.
根据河流中特定站点的卵/幼虫数量,我们使用沿河的卵/幼虫密度分布图外推那一年的总数。
The number of female spawners was then estimated from the mean fecundity, and the number of male spawners and the total number of spawners were estimated from the sex ratio of females to males.
然后根据平均繁殖力估算产卵雌性的数量,根据雌雄性别比估算产卵雄性的数量和产卵者总数。
Estimating initial population sizes
估算初始种群大小
Wei and Yang (32) reported that the average annual catch of Chinese paddlefish in the Yangtze River in the 1970s was approximately 25 metric tons or 676 individuals.
魏和杨(32)报告称,1970年代长江中华匙吻鲟的平均年捕捞量约为25公吨或676个个体。
This is similar to the reported annual catch of 517 individuals for Chinese sturgeon in the 1970s, with an average annual exploitation rate of 23% (50).
这与1970年代中华鲟报告的年捕捞量517个个体相似,平均年利用率为23%(50)。
Both species were commercially harvested in the 1970s, primarily for the spawning stock, with a common catch range in the upper reaches of the Yangtze River.
这两个物种在1970年代都有商业捕捞,主要是为了产卵群体,在长江上游有共同的捕捞范围。
The exploitation rate refers to the ratio of individuals caught to the total number of individuals.
利用率是指捕获个体数与总个体数的比率。
Assuming the same rate for Chinese paddlefish in the 1970s, we estimated that the pre-GD spawning stock size was 2934 individuals.
假设1970年代中华匙吻鲟的利用率相同,我们估计葛洲坝前(pre-GD)的产卵群体大小为2934个个体。
Using a sex ratio of 1:2 (34), we estimated that there were 978 females and 1956 males (text S4.2).
使用1:2的性别比(34),我们估计有978个雌性和1956个雄性(文本S4.2)。
We collected catch data from the 1970s on the total catch and percentage of Yangtze sturgeon in the Yibin, Luzhou, and Hejiang river sections of Sichuan Province.
我们收集了1970年代四川宜宾、泸州和合江河段长江鲟的总捕捞量和百分比数据。
The annual catch of Yangtze sturgeon in the 1970s was estimated to be 9700 kg (text S4.3).
1970年代长江鲟的年捕捞量估计为9700公斤(文本S4.3)。
Assuming that the individuals harvested in this section of the river represent the spawning stock, and using the same exploitation rate as for the Chinese sturgeon (23%), we estimated the spawning stock of the Yangtze sturgeon to be 42,174 kg.
假设在该河段捕捞的个体代表产卵群体,并使用与中华鲟相同的利用率(23%),我们估计长江鲟的产卵群体为42,174公斤。
With an average weight of 10 kg (25), the pre-GD spawning stock was estimated to be 4217 individuals.
以平均体重10公斤(25)计算,葛洲坝前(pre-GD)的产卵群体估计为4217个个体。
Using a sex ratio of 1:2.5, there were 205 females and 3012 males.
使用1:2.5的性别比,有205个雌性和3012个雄性。
Given that mature individuals comprised 6.7% of the total stock (25) and the assumed linear distribution of juvenile density (Fig. 6A), our estimate suggests that there were 58,723 individuals of the recruitment stock (juveniles and subadults) in the Yangtze River, of which 44,042 were in the upper reaches of the Yangtze River (above the GD) and 14,681 in the middle and lower reaches (below the GD).
考虑到成熟的个体占总库存的6.7%(25),以及假设的幼鱼密度线性分布(图6A),我们的估计表明,长江中有58,723个补充群体个体(幼鱼和亚成体),其中44,042个在长江上游(GD以上),14,681个在中下游(GD以下)。
These estimates represent the pre-GD abundance of Yangtze sturgeon.
这些估计代表了葛洲坝前长江鲟的丰富度。
The historical abundance of the Chinese sucker is not well known due to a lack of catch data.
由于缺乏捕捞数据,中华鲴的历史丰富度不太为人所知。
Limited information suggests that from the 1950s to the 1970s, the Chinese sucker became less widespread, and its population notably decreased.
有限的信息表明,从20世纪50年代到70年代,中华鲴的分布范围变小,种群数量明显下降。
The Yibin fisheries cooperatives in Sichuan Province provided the only catch statistics until the 1970s.
四川省宜宾渔业合作社提供了1970年代之前唯一的捕捞统计数据。
We believe that overfishing in the Yibin spawning sites was responsible for the decline.
我们认为,宜宾产卵场地的过度捕捞是导致数量下降的原因。
After the ban on commercial fishing in 1983 and the listing of the Chinese sucker as a second- class protected animal in 1988, individuals of all ages reappeared in the Yangtze, indicating that overfishing did not cause the extinction of the species.
1983年禁止商业捕捞和1988年将中华鲴列为二类保护动物后,所有年龄段的个体在长江重新出现,表明过度捕捞并未导致该物种灭绝。
We used mid- 1970s estimates of pre-GD spawning stock size of 73 individuals (31 females and 42 males) and a juvenile abundance of 8.89 million individuals (text S4.4).
我们使用了1970年代中期葛洲坝前产卵群体大小的估计值,即73个个体(31个雌性和42个雄性)和8.89百万个幼鱼的丰富度(文本S4.4)。
Before 2000, research on C. guichenoti was limited, resulting in a scarcity of relevant data.
2000年之前,对鯮(Coreius guichenoti)的研究有限,导致相关数据匮乏。
Using abundance data from 1997 to 2001 (48), we determined that the initial population size (subadults and adults) below the GD was 265,137 individuals in 1997 (fig. S4A).
利用1997年至2001年的丰富度数据(48),我们确定1997年葛洲坝下游(GD)的初始种群大小(亚成体和成体)为265,137个个体(图S4A)。
Data for the same period above the GD are not available.
GD上游同期的数据不可用。
However, the construction of cascade dams in the Jinsha River has resulted in an increase in research endeavors since 2000.
然而,自2000年以来,金沙江梯级大坝的建设导致研究工作增加。
By analyzing the linear density distributions in Fig. 6C and considering the presence of 570 million eggs/larvae in Panzhihua in 2006 (38), we estimated the total number of eggs/larvae in that year to be 1.981 billion.
通过分析图6C中的线性密度分布,并考虑到2006年攀枝花有5.7亿卵/幼虫(38),我们估计那一年的卵/幼虫总数为19.81亿。
Using an estimate of 22,817 eggs per female (39), we determined that the number of female spawners of C. guichenoti in the upper reaches of the Yangtze River was 86,821 individuals.
使用每雌性22,817个卵的估计值(39),我们确定长江上游鯮(Coreius guichenoti)的产卵雌性数量为86,821个个体。
In 2006, 63,373 males were present, estimated by a sex ratio of 1.37:1, resulting in a total of 150,194 spawners (text S4.5).
2006年,根据1.37:1的性别比,估计有63,373个雄性存在,总共有150,194个产卵者(文本S4.5)。
Dams trigger exponential population declines
大坝引发指数级种群下降
We used the simplified model to reconstruct the population decline of each of the five fishes based on the estimated DICs (Table 1) and initial population sizes.
我们使用简化模型根据估计的DIC值(表1)和初始种群大小重构了这五种鱼类的种群下降。
In the case of the Chinese sturgeon, the males in the spawning stock were more affected by the Yangtze dams than the females and will be the first to go EW.
以中华鲟为例,产卵群体中的雄性受长江大坝的影响比雌性更大,将是首先走向野外灭绝(EW)的。
Figure 8A shows that the Chinese sturgeon spawners declined from 2138 individuals (1033 males) in 1990 to 734 (122) in 2003 to 209 (82) in 2013 to 86 (25) in 2020 to 46 (10) in 2025.
图8A显示,中华鲟的产卵者从1990年的2138个个体(1033个雄性)下降到2003年的734个(122个雄性),2013年的209个(82个雄性),2020年的86个(25个雄性),到2025年的46个(10个雄性)。
Figure 8B shows that the Chinese paddlefish spawners in the Yangtze River consisted of 2934 individuals (1956 males and 978 females) in 1980, which decreased to 40 individuals (27 males and 13 females) in 1993, and only one male in 2005.
图8B显示,长江中华匙吻鲟的产卵者在1980年由2934个个体(1956个雄性和978个雌性)组成,到1993年减少到40个个体(27个雄性和13个雌性),到2005年只剩下一个雄性。
Given that 45 individuals were captured for scientific research between 1982 and 2003 (17), the spawners may have disappeared after 1993, meaning that the Chinese paddlefish stopped breeding and became functionally EX in 1993.
考虑到1982年至2003年之间有45个个体被捕获用于科学研究(17),产卵者可能在1993年之后消失,这意味着中华匙吻鲟停止繁殖,并于1993年实际上灭绝(EX)。
Extinction occurred in 2005 or earlier.
灭绝发生在2005年或更早
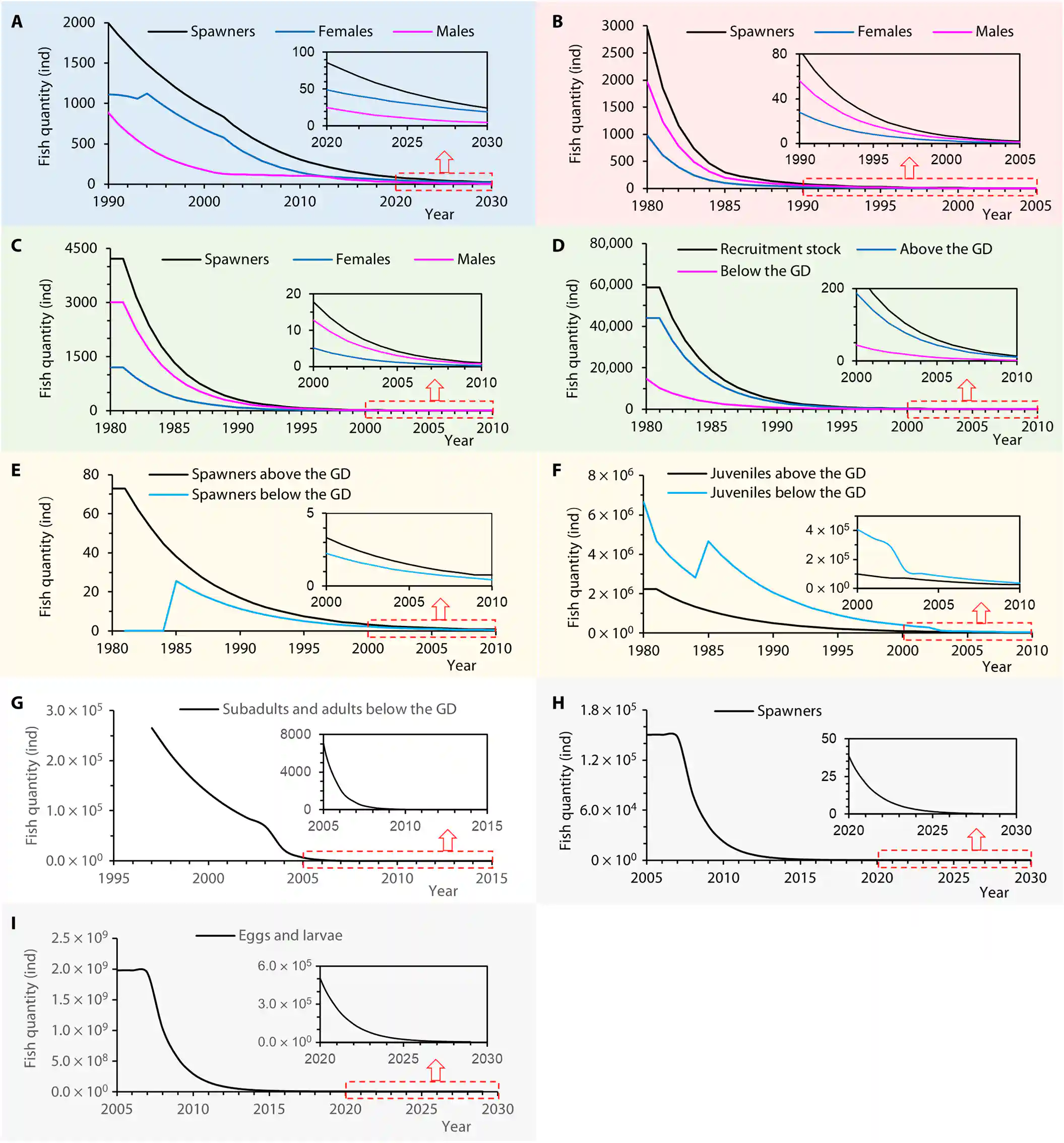
Fig. 8. Population decline processes for the five fishes in the Yangtze River. (A) Chinese sturgeon spawners. (B) Chinese paddlefish spawners. (C and D) Spawning stock and recruitment stock of Yangtze sturgeon. (E and F) Spawners and juveniles of Chinese sucker. (G to I) Total resources (subadults/adults) below the GD, spawners, and eggs/larvae of C. guichenoti. See text S4 for details.
图8. 长江中五种鱼类的种群下降过程。(A) 中华鲟产卵者。(B) 中华匙吻鲟产卵者。(C和D) 长江鲟的产卵群体和补充群体。(E和F) 中华鲴的产卵者和幼鱼。(G至I) GD下游的总资源(亚成体/成体),产卵者和鯮的卵/幼虫。详情请参见文本S4
Figure 8C shows that for the Yangtze sturgeon, the spawners consisted of 4217 individuals (1205 females and 3012 males) in 1980, decreased to 100 (28 and 72) in 1994, and to 18 (5 and 13) in 2000.
图8C显示,对于长江鲟,产卵者在1980年由4217个个体(1205个雌性和3012个雄性)组成,到1994年减少到100个(28个雌性和72个雄性),到2000年减少到18个(5个雌性和13个雄性)。
Given the dispersed breeding manner of Yangtze sturgeon and bycatch mortality, we speculate that breeding activity stopped in 2000 or even earlier, explaining why “no naturally bred juveniles have been found throughout the river since 2000” (51).
考虑到长江鲟分散的繁殖方式和兼捕死亡率,我们推测繁殖活动在2000年或更早就已停止,这解释了为什么“自2000年以来整个河流中没有发现自然繁殖的幼鱼”(51)。
Figure 8D shows that the recruitment stock (juveniles and subadults) was 58,723 individuals (44,042 above the GD and 14,681 below the GD) in 1980, decreased to 1395 (1046 and 244) in 1994, to 248 (186 and 43) in 2000, and lastly to 14 (10 and 2) in 2010.
图8D显示,补充群体(幼鱼和亚成体)在1980年有58,723个个体(GD上游44,042个,下游14,681个),到1994年减少到1395个(1046个上游和244个下游),到2000年减少到248个(186个上游和43个下游),最后到2010年的14个(10个上游和2个下游)。
If we include deaths due to bycatch and fishing for scientific research, all of which occur below the GD, these numbers are even lower.
如果我们包括GD下游由于兼捕和科学研究捕捞导致的死亡,这些数字甚至更低。
This explains why “no wild individuals have been found above the GD since 2010 and below the GD since 1994” (51).
这解释了为什么“自2010年以来GD上游没有发现野生个体,自1994年以来GD下游也没有发现”(51)。
Figures 8 (E and F) shows that the Chinese sucker spawners above the GD decreased from 73 individuals in 1980 to 3 in 2000 to 7 in 2010, while the spawners below the GD decreased from 26 in 1985 to 2 in 2000 to 0 in 2010.
图8(E和F)显示,GD上游的中华鲴产卵者从1980年的73个个体减少到2000年的3个,再到2010年的7个,而GD下游的产卵者从1985年的26个减少到2000年的2个,再到2010年的0个。
Thus, no juveniles have been produced naturally in the Yangtze River since 2010.
因此,自2010年以来,长江中没有自然产生的幼鱼。
The juveniles above the GD decreased from 2.22 million in 1980 to 100,000 in 2000 and to 26,000 in 2010, while the juveniles below the GD decreased from 6.67 million individuals in 1980 to 2.82 million in 1984.
GD上游的幼鱼从1980年的222万个减少到2000年的10万个,再到2010年的2.6万个,而GD下游的幼鱼从1980年的667万个减少到1984年的282万个。
The number of juveniles increased to 4.67 million in 1985 due to the partial natural breeding scale (74%) below the GD, which began in 1985.
由于1985年GD下游开始的部分自然繁殖规模(74%),幼鱼数量在1985年增加到467万个。
After a transient rise in 1985, the juveniles continued to decline to 400,000 in 2000 and to 38,000 in 2010.
在1985年短暂上升之后,幼鱼数量继续下降,到2000年为40万个,到2010年为3.8万个。
Between 1981 and 2003, the population of C. guichenoti below the GD was maintained by eggs/larvae from the upper spawning grounds in the Jinsha River that successfully passed through the turbines at the GD, as the species could not reproduce in the areas below the dam.
在1981年至2003年之间,由于该物种无法在大坝下游的区域繁殖,GD下游的鯮(Coreius guichenoti)种群由来自金沙江上游产卵地并通过GD涡轮机成功通过的卵/幼虫维持。
However, after the construction of the TGD, the eggs/larvae were no longer able to pass through the turbines at the TGD.
然而,在三峡大坝(TGD)建成后,卵/幼虫再也无法通过TGD的涡轮机。
Consequently, the population below the GD disappeared by 2010.
因此,到2010年,GD下游的种群已经消失。
After the impoundment of the cascade dams (Xiangjiaba Dam in 2012 and Xiluodu Dam in 2013), the C. guichenoti population experienced a sharp decline.
在梯级大坝(向家坝于2012年,溪洛渡大坝于2013年)蓄水后,鯮(Coreius guichenoti)的种群经历了急剧下降。
Figure 8G shows that the number of subadults/adults below the GD decreased from 265,137 individuals in 1997 to 70,000 in 2003, when the TGD was put into operation, and further decreased to only 8 in 2011.
图8G显示,GD下游的亚成体/成体数量从1997年的265,137个个体减少到2003年三峡大坝运行时的70,000个,到2011年进一步减少到仅有8个。
Figure 8 (H and I) shows that the spawners decreased from 150,194 individuals in 2006 to 38 in 2020, while eggs/larvae went from 1.981 billion individuals in 2006 to 500,000 in 2020.
图8(H和I)显示,产卵者从2006年的150,194个个体减少到2020年的38个,而卵/幼虫从2006年的1,981亿个个体减少到2020年的500,000个。
Briefly, before the construction of the GD, the five fish species were economically important in the Yangtze River.
简而言之,在葛洲坝建设之前,这五种鱼类在长江中具有重要的经济价值。
However, since the operation of the GD in 1981, the populations of Chinese sturgeon, Chinese paddlefish, Yangtze sturgeon, and Chinese sucker have experienced a substantial decline.
然而,自1981年葛洲坝运行以来,中华鲟、中华匙吻鲟、长江鲟和中华鲴的种群经历了大幅下降。
Although these four species have been legally protected from fishing since 1988, their populations have persistently dwindled.
尽管这四种物种自1988年以来在法律上受到保护,禁止捕捞,但它们的种群数量持续减少。
Similarly, until 2006, C. guichenoti was the primary catch in the upper reaches of the Yangtze River due to the relatively minor impact of the GD and TGD on this species.
同样,直到2006年,由于葛洲坝(GD)和三峡大坝(TGD)对这一物种的影响相对较小,鯮(Coreius guichenoti)是长江上游的主要捕捞对象。
However, the construction of the cascade dams on the Jinsha River, notably the Xiangjiaba and Xiluodu dams, played a fatal role by severing the connection between the spawning stock and the recruitment stocks.
然而,金沙江上的梯级大坝建设,特别是向家坝和溪洛渡大坝,通过切断产卵群体和补充群体之间的联系,发挥了致命的作用。
Therefore, C. guichenoti went from an economically valuable species to an endangered species in a short time.
因此,鯮(Coreius guichenoti)在短时间内从一种经济价值高的物种变成了濒危物种。
As a result, C. guichenoti was officially classified as a second- class national protected species in 2021.
结果,鯮(Coreius guichenoti)在2021年被正式列为二类国家保护物种。
Here, we propose extinction timelines for the five fish species.
在这里,我们为这五种鱼类提出了灭绝时间表。
Specifically, the Chinese sturgeon was considered functionally EX in 2013 and will be EW in 2026.
具体来说,中华鲟在2013年被认为是功能性灭绝(EX),到2026年将彻底野外灭绝(EW)。
The Chinese paddlefish was functionally EX in 1993 and EX in 2005.
中华匙吻鲟在1993年功能性灭绝,2005年彻底灭绝。
In addition, the Yangtze sturgeon and the Chinese sucker were EW in 2010, while the C. guichenoti will be EW by 2030.
此外,长江鲟和中华鲴在2010年已经野外灭绝,而鯮(Coreius guichenoti)到2030年将野外灭绝。
Migration patterns have varying effects on population declines.
迁徙模式对种群下降有不同的影响。
Among diadromous fishes, freshwater amphidromous fishes (e.g., Chinese paddlefish) are more affected by dams than anadromous fishes (e.g., Chinese sturgeon) because only a small proportion of the anadromous population develops into the spawning stock that enters the river to breed each year, while a large proportion of the recruitment stock remains in the sea.
在洄游性鱼类中,淡水两栖洄游性鱼类(例如中华匙吻鲟)比溯河洄游性鱼类(例如中华鲟)受大坝的影响更大,因为只有一小部分溯河洄游性种群每年发育成为进入河流繁殖的产卵群体,而大部分补充群体留在海中。
Therefore, even when breeding in the river ceases, the migratory phenomenon can persist for several years or even decades, creating an illusion of species population health or sustainability based on the presence of invalid stocks.
因此,即使河流中的繁殖停止,洄游现象仍可能持续数年甚至数十年,这基于无效种群的存在,创造了一种物种种群健康或可持续性的错觉。
Among riverine potamodromous species, the fish that cannot reproduce below the dam (e.g., Yangtze sturgeon and C. guichenoti) are more severely affected by dams than species that can reproduce below the dam (e.g., Chinese sucker), suggesting lower life history plasticity.
在河流性河川洄游性物种中,不能在大坝下游繁殖的鱼类(例如长江鲟和鯮)比能在大坝下游繁殖的物种(例如中华鲴)受大坝的影响更严重,这表明它们的生活史可塑性较低。
DISCUSSION
讨论
Scientific misjudgments underpinning the FRPs
支撑鱼类救援计划(FRPs)的科学误判
After four decades of implementation of the FRPs, the five prominent fish species are facing imminent extinction.
在实施鱼类救援计划(FRPs)四十年后,这五种著名的鱼类正面临即将到来的灭绝。
China has undertaken conservation efforts since 2015, with two action plans specifically aimed at conserving the Chinese sturgeon and the Yangtze sturgeon.
自2015年以来,中国已经开展了保护工作,有两个行动计划专门针对保护中华鲟和长江鲟。
However, these plans do not adequately address the negative impacts of dams and lack targeted measures (51, 52).
然而,这些计划没有充分解决大坝的负面影响,也缺乏针对性的措施(51, 52)。
In addition, China imposed a 10- year fishing ban on the Yangtze River in 2020, followed by enacting the Yangtze River Protection Law in 2021.
此外,中国在2020年对长江实施了为期10年的禁渔令,随后在2021年颁布了《长江保护法》。
This legislation designates 13 aquatic species, including the five fishes studied here, as CR (53).
这项立法将13种水生物种,包括这里研究的五种鱼类,指定为极危物种(CR)(53)。
However, we believe that these measures will not be sufficient to recover the five fishes in the Yangtze River because it is imperative to address the dangerous situation created by the Yangtze dams.
然而,我们认为这些措施不足以恢复长江中的五种鱼类,因为必须解决长江大坝所造成的危险局面。
Our findings suggest that the FRPs for the Yangtze dams are scientifically based on unreliable judgments, resulting in a flawed fish conservation policy.
我们的发现表明,长江大坝的鱼类救援计划(FRPs)是基于不可靠的科学判断,导致了一个有缺陷的鱼类保护政策。
Here, we present six major misjudgments that underlie China’s FRPs (text S5).
在这里,我们提出了支撑中国FRPs的六个主要误判(文本S5)。
The first misjudgment concerns the assertion that overfishing is the primary cause of the population collapse of the five fishes.
第一个误判是断言过度捕捞是这五种鱼类种群崩溃的主要原因。
Our studies have shown that the Yangtze dams were primarily responsible for the decline of the Chinese sturgeon population (13, 30, 54, 55).
我们的研究表明,长江大坝是中华鲟种群数量下降的主要原因(13, 30, 54, 55)。
However, IHB ichthyologists have chosen to disregard the failure of the FRPs and instead attribute the survival crisis of the five fishes, especially the Chinese sturgeon, to overfishing (56, 57).
然而,中国科学院水生生物研究所(IHB)的鱼类学家选择忽视FRPs的失败,而是将这五种鱼类的生存危机,特别是中华鲟,归因于过度捕捞(56, 57)。
Our findings indicate that the Chinese sturgeon has not been overfished, as the average annual exploitation rate from 1972 to 2017 was 0.61%, with a peak of 3.6% in 1981, which is below the 5% threshold for overfishing (Fig. 9).
我们的研究结果表明,中华鲟并未被过度捕捞,因为从1972年到2017年的平均年利用率为0.61%,在1981年达到高峰3.6%,这低于5%的过度捕捞阈值(图9)。
Our research has consistently found no evidence to support overfishing as the primary factor contributing to the survival crisis of the five fish species.
我们的研究一直未发现支持过度捕捞是导致这五种鱼类生存危机的主要因素的证据。
Instead, we highlight that the Yangtze dams, not overfishing, have disrupted the life cycles of the five fishes by blocking migration and hindering reproductive success.
相反,我们强调的是长江大坝,而不是过度捕捞,通过阻碍迁徙和妨碍繁殖成功,扰乱了这五种鱼类的生命周期。
Moreover, the lack of targeted measures has led to the emergence of large invalid stocks.
此外,缺乏针对性的措施导致了大量无效种群的出现。
As a result, merely banning fishing without addressing the effects of dams is insufficient to save these five fish species.
因此,仅仅禁止捕鱼而不解决大坝的影响,是不足以拯救这五种鱼类的。
The second misjudgment involves the claim that “breeding below the dam” indicates species conservation success.
第二个误判涉及到“大坝下游的繁殖”表明物种保护成功的主张
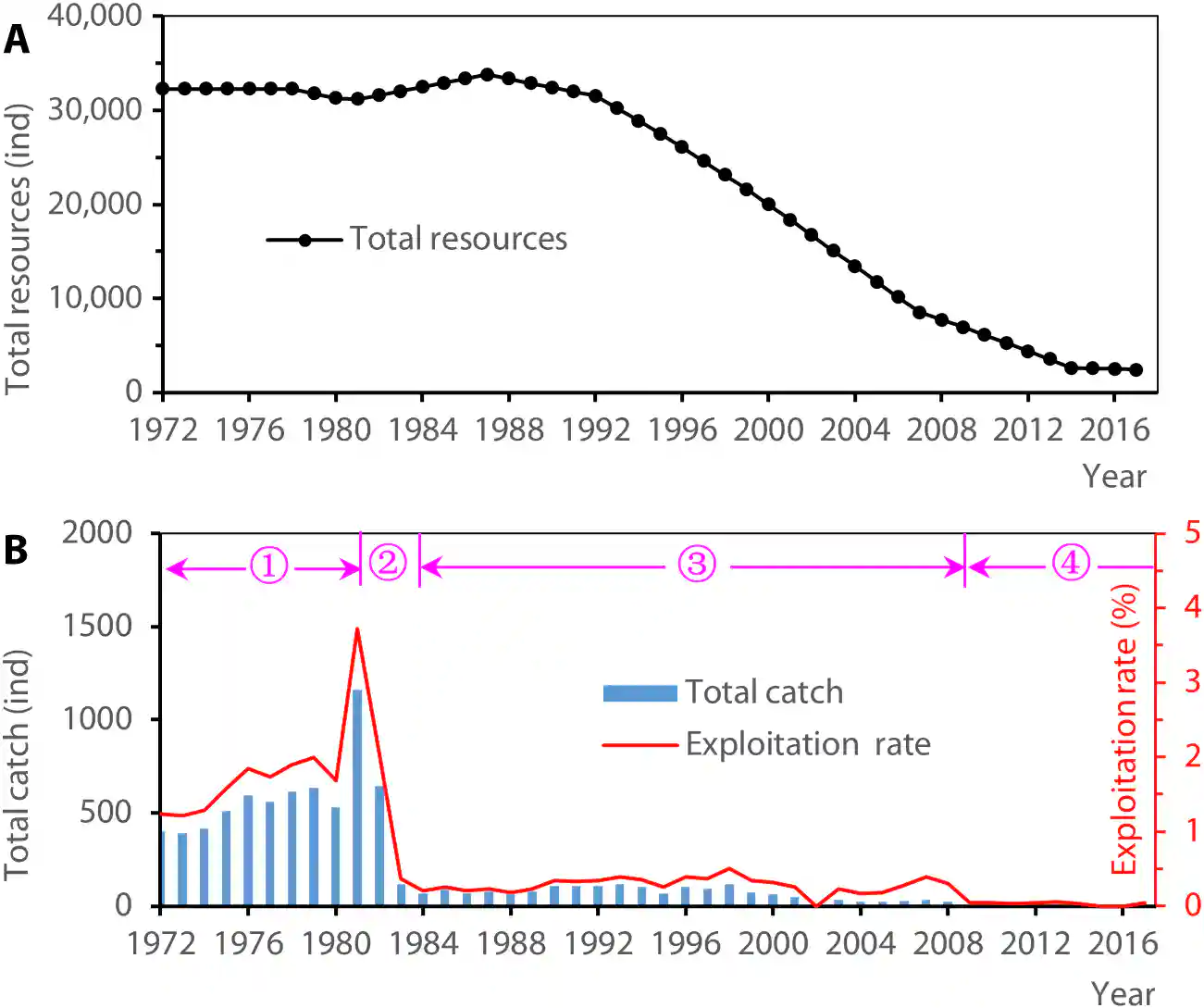
Fig. 9. Estimated exploitation rate of the Chinese sturgeon. (A) Total population size. (B) Total catch number and annual exploitation rate. ① Steady commercial fishing (pre-GD and before 1981), with an average exploitation rate of 1.6%. ② Initial stage of post-GD (1981–1982), with an average exploitation rate of 2.8%. ③ Fishing restricted to scientific purposes (1983–2008), with an average exploitation rate of 0.3%. ④ Comprehensive fishing ban (2009–present), with an average exploitation rate of 0.04%. An exploitation rate of 5% is a threshold for overfishing of the Chinese sturgeon. See text S5 for details.
图9. 中华鲟的估计开发率。(A) 总人口大小。(B) 总捕捞数量和年开发率。① 稳定的商业捕捞(GD前和1981年前),平均开发率为1.6%。② GD后初期(1981–1982年),平均开发率为2.8%。③ 限于科学研究目的的捕捞(1983–2008年),平均开发率为0.3%。④ 全面禁渔(2009年至今),平均开发率为0.04%。5%的开发率是中华鲟过度捕捞的阈值。详情请参见文本S5
In January 1981, as the river closure of the GD approached, the pending fish rescue measures triggered a major debate on how to protect migratory fish in China.
1981年1月,随着葛洲坝(GD)的河流封闭临近,即将实施的鱼类救援措施引发了一场关于如何保护中国迁徙性鱼类的重大辩论。
A particular concern arose regarding the Chinese sturgeon’s ability to continue breeding after the river was dammed.
人们特别担心中华鲟在河流被大坝截断后是否还能继续繁殖。
The IHB reported that its scientists caught eggs, larvae, and mature individuals below the GD in the fall of 1981 and 1982, suggesting that Chinese sturgeon could adapt to the dam-modified environment and continue to reproduce.
中国科学院水生生物研究所(IHB)报告称,其科学家在1981年和1982年秋季在GD下游捕获了卵、幼虫和成熟的个体,这表明中华鲟能够适应大坝改造后的环境并继续繁殖。
Therefore, the IHB recommended restocking as the primary measure instead of fish passage.
因此,IHB建议将人工增殖放流作为主要措施,而不是建立鱼道。
The discovery of “mature” Chinese paddlefish and Chinese sucker below the GD implies that their wild populations would not be extirpated (58–60).
在GD下游发现“成熟”的中华匙吻鲟和中华鲴,意味着它们的野生种群不会被根除(58-60)。
However, our findings indicate that the Chinese paddlefish population declined rapidly and became EX after the construction of the GD because the juveniles were unable to return to their spawning grounds after migrating to the sea.
然而,我们的发现表明,中华匙吻鲟的种群在GD建成后迅速下降并成为灭绝(EX),因为幼鱼在迁徙到海里后无法返回它们的产卵场。
In addition, our findings show that breeding below the dam does not prevent extinction and is not as important as some ichthyologists expect.
此外,我们的发现表明,大坝下游的繁殖并不能阻止灭绝,也不像一些鱼类学家预期的那样重要。
The spawning stocks of Chinese sturgeon and Chinese sucker experienced notable declines, with a large portion becoming invalid stock each year.
中华鲟和中华鲴的产卵群体经历了显著的下降,每年都有很大一部分变成了无效种群。
Here, we emphasize that previous studies have used the presence of invalid stocks to suggest the existence of a viable and sustainable population, but this conceals the inevitable decline of these populations toward extinction (14, 58–60).
在这里,我们强调以前的研究使用无效种群的存在来表明有一个可行和可持续的种群,但这掩盖了这些种群不可避免地走向灭绝的下降趋势(14, 58-60)。
The third misjudgment is regarding the claim that restocking is a viable strategy for mitigating the adverse impact of dam-induced fish depletion.
第三个误判涉及到人工增殖放流是缓解大坝引起的鱼类枯竭不利影响的可行策略的主张。
Restocking involves introducing artificially bred fish into existing populations to replenish diminished spawning biomass.
人工增殖放流涉及将人工繁殖的鱼引入现有种群,以补充减少的产卵生物量。
In 1982, the IHB suggested that restocking could effectively offset the decline of the Chinese sturgeon population caused by the GD (14).
1982年,IHB建议人工增殖放流可以有效抵消葛洲坝(GD)造成的中华鲟种群数量下降(14)。
As a result, the Institute of Chinese Sturgeon was established in 1983 with a specific focus on researching artificial breeding and release technology.
因此,1983年成立了中华鲟研究所,专门研究人工繁殖和放流技术。
In addition, many professional institutions have researched the artificial propagation and release of Chinese sturgeon and other migratory species, making restocking a popular solution to China’s dam impact on fish species.
此外,许多专业机构已经研究了中华鲟和其他迁徙性物种的人工繁殖和放流,使人工增殖放流成为中国应对大坝对鱼类种群影响的流行解决方案。
Since 1983, more than seven million Chinese sturgeon of various sizes have been released into the Yangtze River (13, 61).
自1983年以来,已有超过七百万条各种大小的中华鲟被放流到长江中(13, 61)。
However, there is currently no strong evidence that any of the released individuals have returned to the Yangtze River and engaged in reproductive activities.
然而,目前没有强有力的证据表明任何放流的个体已经返回长江并参与繁殖活动。
Therefore, we have argued that the restocking efforts for the Chinese sturgeon have been unsuccessful (62), but not because of the alleged insufficiency in the quantity and size of released fish, as some ichthyologists have suggested (57, 61, 63).
因此,我们认为中华鲟的人工增殖放流努力是不成功的(62),但并不是因为一些鱼类学家所建议的放流鱼的数量和大小不足(57, 61, 63)。
Restocking may be a better solution to overfishing than addressing the dam barrier effect.
与解决大坝障碍效应相比,人工增殖放流可能是解决过度捕捞问题的一个更好方案。
The study’s findings suggest that the success of FRPs is considerably dependent on the preservation of the fish life cycle, specifically the facilitation of reproduction in natural habitats.
该研究的发现表明,鱼类救援计划(FRPs)的成功在很大程度上依赖于保护鱼的生活周期,特别是促进在自然栖息地的繁殖。
The fourth misjudgment concerns the claim that replacing mainstream habitats with tributaries can protect flagship species.
第四个误判涉及到用支流替代主流生境可以保护旗舰物种的主张。
Since 2000, Chinese ichthyologists have proposed a fish conservation strategy called “replacing mainstream habitats with tributaries” to mitigate the negative impacts of cascade dams on the mainstream of the Jinsha River.
自2000年以来,中国的鱼类学家提出了一种名为“用支流替代主流生境”的鱼类保护策略,以减轻梯级大坝对金沙江主流的负面影响。
This involves designating key tributaries as dam- free, free- flowing rivers by either prohibiting dams or dismantling small dams.
这包括通过禁止建造大坝或拆除小型大坝,将关键支流指定为无坝、自由流动的河流。
Cao (64), an ichthyologist at the IHB, noted that if dams block migrating fish in the Jinsha mainstream, then they will swim back to the tributaries to reproduce.
中国科学院水生生物研究所(IHB)的鱼类学家曹(64)指出,如果大坝阻挡了金沙江主流的迁徙性鱼类,那么它们将游回支流进行繁殖。
This approach was developed by the TGD- FRP in the 1990s.
这种方法在20世纪90年代由三峡大坝鱼类救援计划(TGD-FRP)发展。
In the TGD reservoir area, 44 endemic species were observed, 29 of which were also found in the Chishui River (11).
在三峡大坝库区,观察到了44种特有物种,其中29种也出现在赤水河中(11)。
The Chishui River has been identified as a potential alternative habitat for fish species affected by the TGD (fig. S5).
赤水河已被确定为受三峡大坝影响的鱼类物种的潜在替代栖息地(图S5)。
However, this approach cannot save the five flagship fishes or modify their life cycle because they primarily migrate in the mainstream of the Yangtze River and not in its tributaries.
然而,这种方法无法拯救这五种旗舰鱼类或改变它们的生活周期,因为它们主要在长江主流中迁徙,而不是在其支流中。
The fifth misjudgment pertains to the assertion that the FRPs were backed by solid science.
第五个误判涉及到FRPs得到坚实科学支持的断言。
The selection of fish rescue targets for dams is critical.
为大坝选择鱼类救援目标至关重要。
However, the prioritization of these targets is flawed due to knowledge gaps in fish migration taxonomy and life cycle integrity.
然而,由于鱼类迁徙分类学和生活周期完整性的知识空白,这些目标的优先级排序存在缺陷。
The GD- FRP has focused exclusively on the Chinese sturgeon, ignoring the Chinese paddlefish, the Yangtze sturgeon, and the Chinese sucker.
葛洲坝鱼类救援计划(GD-FRP)专门关注中华鲟,忽视了中华匙吻鲟、长江鲟和中华鲴。
Our findings indicate that the priority of the GD- FRP should be determined by the severity of the impact, with the Chinese paddlefish ranking highest (β = 0.63 to 0.78), followed by the Yangtze sturgeon (0.75), Chinese sucker (0.74 to 0.85), Chinese sturgeon (0.88 to 0.93), and C. guichenoti (0.88 to 0.98).
我们的发现表明,GD-FRP的优先级应根据影响的严重程度来确定,中华匙吻鲟的优先级最高(β = 0.63至0.78),其次是长江鲟(0.75),中华鲴(0.74至0.85),中华鲟(0.88至0.93),以及鯮(Coreius guichenoti)(0.88至0.98)。
The Chinese paddlefish, Yangtze sturgeon, and Chinese sucker are more threatened by the GD than the Chinese sturgeon.
中华匙吻鲟、长江鲟和中华鲴比中华鲟更受到GD的威胁。
Unfortunately, they were not prioritized in the GD- FRP, resulting in their extinction before the TGD- FRP was implemented.
不幸的是,它们在GD-FRP中没有被优先考虑,导致在TGD-FRP实施之前就已经灭绝。
In the mid- 1990s, a national rare fish reserve was established in the Hejiang- Pingshan section of the upper reaches of the Yangtze River as a key measure of the TGD- FRP.
在20世纪90年代中期,作为三峡大坝鱼类救援计划(TGD-FRP)的关键措施,在长江上游的合江-屏山段建立了一个国家级珍稀鱼类保护区。
However, the barrier effect of the GD had resulted in the extinction or near extinction of protected species such as the Chinese paddlefish, Yangtze sturgeon, and Chinese sucker before the implementation of the TGD- FRP, so this reserve exists in name only.
然而,在TGD-FRP实施之前,GD的阻碍效应已经导致了像中华匙吻鲟、长江鲟和中华鲴这样的保护物种的灭绝或接近灭绝,因此这个保护区只是名义上的存在。
Our findings contradict the purported success and scientific foundation of the GD- and TGP- FRPs (56), indicating the opposite.
我们的发现与声称的GD和TGP-FRPs(56)的成功和科学基础相矛盾,表明情况正好相反。
The sixth misjudgment concerns the assertion that fishways are unnecessary in dams.
第六个误判涉及到大坝中不需要鱼道的断言。
The 1982 GD- FRP suggested that fishways were not needed for the Chinese sturgeon (14).
1982年的GD-FRP建议中华鲟不需要鱼道(14)。
The TGD, built in 1993, followed this idea and did not include fishways.
1993年建成的三峡大坝(TGD)遵循了这一理念,没有包括鱼道。
Despite a fish passage policy implemented by the Chinese government after 2000, 10 dams built since then upstream of the TGD still lack fishways.
尽管中国政府在2000年后实施了鱼类通道政策,但自那时起在TGD上游建造的10座大坝仍然没有鱼道。
The underlying reason is that the IHB’s fish rescue strategy, which lacks fishways for the GD and TGD, has received considerable scientific and technical awards and recognition.
其根本原因是,中国科学院水生生物研究所(IHB)的鱼类救援策略,即GD和TGD不需要鱼道,已经获得了相当的科学和技术奖项及认可。
In addition, it has been widely supported by the Chinese hydropower industry as an exemplary approach to fish conservation in the hydropower sector.
此外,它得到了中国水电行业的广泛支持,被视为水电领域鱼类保护的典范方法。
The results of this study indicate that fish passage is the most favorable approach to mitigate the barrier effect of dams on migratory fish.
本研究的结果表明,鱼类通道是减轻大坝对迁徙性鱼类障碍效应的最有利方法。
Although doubts have been raised regarding the efficacy of fish passage (65), we argue that the implementation of fish passage is imperative in cases where the dam poses a threat to migratory fish populations.
尽管对鱼道的有效性提出了疑问(65),但我们认为,在大坝对迁徙性鱼类种群构成威胁的情况下,实施鱼道是必要的。
Consequently, we advocate the restoration of upstream and downstream connectivity in the Yangtze River.
因此,我们主张恢复长江的上下游连通性。
In particular, China’s fish passage policy needs improvement, as fishways built after 2000 only consider the upstream migration of the spawning stock, ignoring the downstream migration of the recruitment stock (larvae, juveniles, and post- spawners) (66).
特别是,中国的鱼道政策需要改进,因为2000年后建造的鱼道只考虑了产卵群体的上游迁移,忽视了补充群体(幼虫、幼鱼和产卵后个体)的下游迁移(66)。
Reforming China’s FRP for dams
改革中国的大坝鱼类救援计划(FRP)
Most of the world’s rivers are dammed, including new dams under construction.
世界上大部分河流都建有大坝,包括正在建设中的新大坝。
Protecting migratory fish from the threats posed by dams is a common challenge worldwide, including in China.
保护迁徙性鱼类免受大坝威胁是一个全球性的共同挑战,包括在中国。
However, despite China’s efforts to protect the Yangtze River, the negative impacts of dams on migratory fish have received limited attention due to the complex nature of the issue.
然而,尽管中国努力保护长江,但由于问题的复杂性,大坝对迁徙性鱼类的负面影响受到了有限的关注。
To protect fish species, it is critical to learn from four decades of FRPs and focus on reforming China’s dam- related FRP.
为了保护鱼类物种,从四十年的鱼类救援计划(FRPs)中学习,并专注于改革中国与大坝相关的FRP至关重要。
China needs to recognize scientific misjudgments to redirect research toward innovation.
中国需要认识到科学上的误判,并将研究引向创新。
The failure of the FRPs serves as a stark reminder of the need to conduct a comprehensive examination of the reasons behind the ineffectiveness of China’s fish conservation strategies and their purported “scientific results.”
FRPs的失败是一个严峻的提醒,需要对中国鱼类保护策略及其所谓的“科学成果”无效的原因进行全面审查。
China has invested heavily in scientific research on dam- related fish conservation over the past 40 years.
过去40年,中国在与大坝相关的鱼类保护科学研究上投入巨大。
However, many of these scientific research efforts have proven inadequate to support the conservation of flagship fish species, primarily due to the misguided direction of basic research.
然而,由于基础研究的方向错误,这些科学研究工作证明不足以支持旗舰鱼种的保护。
To reformulate FRPs for the Yangtze dams, China must prioritize correcting misconceptions about past FRPs, including the six major misjudgments mentioned above.
为了重新制定长江大坝的FRPs,中国必须优先纠正对过去FRPs的误解,包括上述的六个主要误判。
China needs to strengthen oversight of dam owners to clarify their obligations to protect fish.
中国需要加强对大坝所有者的监管,明确他们保护鱼类的义务。
These five fishes were the main targets of the FRPs for the Yangtze dams. Who exactly is responsible for saving these fishes?
这五种鱼是长江大坝FRPs的主要目标。究竟是谁负责拯救这些鱼类?
According to China’s environmental protection laws, the hydropower companies (dam owners) are responsible for protecting the affected fish.
根据中国的环保法律,水电公司(大坝所有者)负责保护受影响的鱼类。
However, because of the complexity of factors affecting fish, such as overfishing, water pollution, and navigation, the responsibility of hydropower companies has often not been recognized, and they have been eager to fund authoritative institutions to absolve themselves of blame, rather than seeking ways to implement their responsibility to protect fish.
然而,由于影响鱼类的因素复杂,如过度捕捞、水污染和航运,水电公司的责任往往没有得到认可,他们急于资助权威机构以免除自己的责任,而不是寻找方法来履行他们保护鱼类的责任。
We suggest that China strengthen the supervision of hydropower companies and incorporate fish conservation in the strict rating criteria so that the companies treat fish conservation as a true concern of the companies themselves.
我们建议中国加强对水电公司的监管,并将鱼类保护纳入严格的评级标准,以便公司将鱼类保护视为公司自身的真正关切。
China needs to use adaptive management to handle intricate ecological projects effectively.
中国需要使用适应性管理来有效地处理复杂的生态项目。
This involves a comprehensive set of procedures and institutional arrangements, including an “implementation, monitoring, evaluation, and improvement” cycle to continuously review and improve fish rescue strategies and countermeasures.
这涉及一套全面的程序和制度安排,包括一个“实施、监测、评估和改进”循环,以不断审查和改进鱼类救援策略和对策。
Since 1995, China has established an ecological and environmental monitoring network for the TGD (20), which has proven to be useless for conserving these flagship fishes.
自1995年以来,中国为三峡大坝(TGD)建立了一个生态和环境监测网络(20),事实证明这对保护这些旗舰鱼类毫无用处。
To solve this problem, we propose to develop and implement an ecological operation plan for the Yangtze cascade dams to restore the reproductive hydrological conditions of the affected migratory fish species based on an adaptive management system and the monitoring network to constantly improve the operation program.
为了解决这个问题,我们提议开发和实施一个长江梯级大坝的生态运营计划,以恢复受影响的迁徙性鱼类的繁殖水文条件,该计划基于适应性管理系统和监测网络,不断改进运营计划。
China also needs to improve international cooperation for innovation and knowledge sharing.
中国还需要加强国际合作,促进创新和知识共享。
Conflicts between dams and migratory fish are common in rivers worldwide (1–9).
大坝和迁徙性鱼类之间的冲突在全球河流中很常见(1-9)。
Therefore, international cooperation is urgently needed to address these challenges and share experience in maintaining the balance between fish conservation and dam management.
因此,迫切需要国际合作来应对这些挑战,并分享在保护鱼类和大坝管理之间保持平衡的经验。
In addition, international rivers that are undergoing hydropower development, such as the Amazon, Congo, and Mekong, require cooperative efforts to conserve migratory fish populations effectively.
此外,正在进行水电开发的国际河流,如亚马逊河、刚果河和湄公河,需要合作努力以有效保护迁徙性鱼类种群。
International cooperation is urgently needed for China to enhance fish passage and ecological operation techniques (66, 67).
中国迫切需要国际合作来加强鱼类通道和生态运营技术(66, 67)。
The five fishes: Hope or concern?
这五种鱼:希望还是担忧?
Now, the five fish species have already lost their best chance for recovery.
现在,这五种鱼类已经失去了恢复的最佳机会。
The destruction of spawning grounds, feeding grounds, and migration corridors can harm migratory fish and cause population extinction with the loss of any one of these three habitat elements (68).
产卵场、觅食场和迁徙通道的破坏可以伤害迁徙性鱼类,并导致任何一个这三种栖息地要素的丧失都可能导致种群灭绝(68)。
Unfortunately, all three habitats for the five species have been destroyed by the Yangtze dams.
不幸的是,这五种物种的三种栖息地都被长江大坝破坏了。
The Chinese paddlefish were EX in 2005, while the remaining four species are on the verge of extinction, with only hatchery-bred individuals surviving.
中华匙吻鲟在2005年已经灭绝(EX),而剩下的四个物种正濒临灭绝,只有人工繁殖的个体存活。
Therefore, it is imperative to mitigate the barrier effects of dams and supplement natural populations through restocking and rewilding.
因此,迫切需要减轻大坝的阻碍效应,并通过人工增殖放流和重新野化来补充自然种群。
How can the barrier effect of dams be mitigated?
如何减轻大坝的阻碍效应?
There are three potential solutions to address this issue: removing the dams, constructing fish passages, or implementing the ecological operation of the dams.
解决这个问题有三个潜在的解决方案:拆除大坝、建造鱼道或实施大坝的生态运行。
However, the feasibility of removing the Yangtze dams in China remains impractical despite the growing prevalence of dam removal initiatives in the United States and Europe (69).
然而,尽管美国和欧洲拆除大坝的倡议越来越普遍(69),但在中国拆除长江大坝的可行性仍然不切实际。
The construction of fishways for these dams presents huge technical obstacles that make success unlikely in the short term.
对于这些大坝建造鱼道面临着巨大的技术障碍,这使得短期内成功的可能性不大。
Consequently, the sole viable alternative lies in the ecological management of the cascade dams.
因此,唯一可行的替代方案在于梯级大坝的生态管理。
Hence, our recommendations for the five fish species must be based on the existing circumstances.
因此,我们对这五种鱼类的建议必须基于现有的情况。
To save the Chinese sturgeon, it is imperative to reduce the water temperature in the spawning area below the GD by 2.7°C from October 22 to November 15 (30).
为了拯救中华鲟,必须在10月22日至11月15日期间将葛洲坝下游产卵区的水温降低2.7°C(30)。
To achieve this goal, we propose to integrate the ecological operation of the cascade dams with local cooling techniques to maintain an appropriate water temperature range (18° to 20°C).
为了实现这一目标,我们提议将梯级大坝的生态运行与当地的冷却技术相结合,以维持适当的水温范围(18°至20°C)。
In addition, as a complementary measure, we recommend improving the release efficiency of the Chinese sturgeon restocking program so that the species can return to the Yangtze River and reproduce successfully.
此外,作为补充措施,我们建议提高中华鲟人工增殖放流计划的释放效率,以便该物种能够返回长江并成功繁殖。
The Yangtze sturgeon, Chinese sucker, and C. guichenoti are riverine potamodromous fish species that primarily breed in the upper Yangtze River, including a 400- km section from Yibin to Chongqing between the Xiangjiaba Dam and the end of the TGD reservoir, where some free- flowing features still exist.
长江鲟、中华鲴和鯮是河流性河川洄游性鱼类,主要在长江上游繁殖,包括向家坝大坝和三峡大坝水库末端之间的宜宾至重庆400公里河段,这里仍保留一些自由流动的特征。
This study shows that several individuals from the three species were able to complete their life cycles in the 400- km section.
这项研究表明,这三个物种的一些个体能够在400公里的河段中完成它们的生活周期。
It is imperative to investigate the life history plasticity of these species and develop strategies to mitigate the barrier effects of the dams.
迫切需要研究这些物种的生活史可塑性,并制定策略以减轻大坝的阻碍效应。
Conservation efforts for these species could be enhanced by promoting the successful completion of their life cycles, particularly their reproductive behavior, within the section.
通过促进这些物种在该河段内成功完成它们的生活周期,特别是它们的繁殖行为,可以加强对这些物种的保护努力。
METHODS Fish migration taxonomy
方法:鱼类迁徙分类学
The study of modern fish migration has a history of over 100 years since the publication of Meek’s book in 1916 (70).
自1916年米克(Meek)的书籍出版以来,现代鱼类迁徙的研究已有100多年的历史(70)。
Taxonomic nomenclature and definitions were widely accepted until Myers’ work in 1949 (71).
直到1949年迈尔斯(Myers)的工作之前,分类命名和定义被广泛接受(71)。
Later, McDonald and others (72–77) further improved the definitions, evolution, and application of fish migration classification.
后来,麦克唐纳(McDonald)和其他人(72-77)进一步完善了鱼类迁徙分类的定义、演变和应用。
Fish migration taxonomy was based on the marked differences in salt content between freshwater and saltwater.
鱼类迁徙分类学是基于淡水和咸水之间盐分含量的显著差异。
Diadromous fish migrate between freshwater and saltwater, potamodromous fish migrate entirely within freshwater, and oceanodromous fish migrate entirely within seawater (71, 72).
洄游性鱼类在淡水和咸水之间迁徙,河川洄游性鱼类完全在淡水中迁徙,而海洋洄游性鱼类完全在海水中迁徙(71, 72)。
The life cycle of a migratory fish includes various growth stages related to its migration, from fertilized egg to larva, juvenile, subadult, adult, spawner, and post-spawner.
迁徙性鱼类的生命周期包括与其迁徙相关的各种生长阶段,从受精卵到幼虫、幼鱼、亚成体、成体、产卵者和产卵后个体。
This life cycle can vary greatly depending on species, habitat, and other environmental factors (72).
这个生命周期可以根据物种、栖息地和其他环境因素而大不相同(72)。
However, definitions of migration classification are assigned based solely on the most distinctive migration in the specific life stage of a given fish (e.g., reproductive migration) and do not take into account other migrations that occur throughout the life cycle of migratory fish.
然而,迁徙分类的定义仅基于特定鱼类(例如,繁殖迁徙)的特定生活阶段中最显著的迁徙,并不考虑迁徙性鱼类整个生命周期中发生的其他迁徙。
Therefore, it is impossible to understand the entire life cycle of fish by relying on the classification of migration alone.
因此,仅依靠迁徙分类是无法理解鱼类整个生命周期的。
Here, we propose a three-tiered (category- type- subtype) framework for fish migration taxonomy (Fig. 2).
在这里,我们提出了一个三层级(类别-类型-亚型)的鱼类迁徙分类框架(图2)。
In this framework, migratory fishes are divided into three categories—diadromy, potamodromy, and oceanodromy.
在这个框架中,迁徙性鱼类被分为三个类别—洄游性、河川洄游性和海洋洄游性。
Diadromy is subdivided into three types, namely, anadromy, catadromy, and amphidromy, of which the amphidromy is further subdivided into two subtypes based on the spawning environment, namely, freshwater amphidromy and marine amphidromy.
洄游性进一步细分为三种类型,即溯河洄游性、降河洄游性和两栖洄游性,其中两栖洄游性根据产卵环境进一步细分为两个亚型,即淡水两栖洄游性和海洋两栖洄游性。
Previous studies have defined the three types (anadromy, catadromy, and amphidromy) and the two subtypes (freshwater amphidromy and marine amphidromy) of diadromy (71, 73, 74).
以前的研究已经定义了洄游性的三种类型(溯河洄游性、降河洄游性和两栖洄游性)和两种亚型(淡水两栖洄游性和海洋两栖洄游性)(71, 73, 74)。
Here, we propose to subdivide potamodromy, which is largely dam related and less studied, into two types: riverine potamodromy and lacustrine potamodromy.
在这里,我们提议将与大坝关系较大且研究较少的河川洄游性进一步细分为两种类型:河流性河川洄游性和湖泊性河川洄游性。
Definitions of migration patterns
迁徙模式的定义
Diadromy was first used by Myers in 1949 to describe “truly migratory fishes that migrate between the sea and freshwater” (71).
洄游性这个术语最初由迈尔斯在1949年使用,用来描述“真正在海洋和淡水之间迁徙的鱼类”(71)。
Diadromy is the earliest described migration pattern, and diadromous fish are the most well-studied category of migratory fish.
洄游性是最早描述的迁徙模式,洄游性鱼类也是迁徙性鱼类中最研究得最充分的类别。
McDowall (74, 75) refined this definition to stipulate that diadromous migrations must occur regularly, be physiologically mediated movements between two biomes, occur at predictable times, occur during a characteristic life history phase in each species, involve most individuals in a given population, be typically obligatory, and involve two reciprocal migrations, from freshwater to the sea and the reverse.
麦克道尔(McDowall)(74, 75)对这一定义进行了完善,规定洄游性迁徙必须定期发生,是两个生物群落之间生理介导的运动,发生在可预测的时间,发生在每个物种特有的生命周期阶段,涉及给定种群中的大多数个体,通常是强制性的,并涉及两个相反方向的迁徙,从淡水到海洋以及相反方向。
Myers (71) defined anadromy as “diadromous fishes that spend most of their lives in the sea and migrate to freshwater to breed.”
迈尔斯(Myers)(71)将溯河洄游性定义为“大部分生命时间在海中度过,迁徙到淡水中繁殖的洄游性鱼类”。
McDowall (74, 75) refined the definition as follows: “diadromous fishes in which most feeding and growth are at sea before the migration of fully grown, adult fish into freshwater to reproduce; either there is no subsequent feeding in freshwater, or any feeding is accompanied by little somatic growth; the principal feeding and growing biome (the sea) differs from the reproductive biome (freshwater).”
麦克道尔(McDowall)(74, 75)对定义进行了如下完善:“在完全长大的成年鱼迁徙到淡水中繁殖之前,大多数觅食和生长在海中进行的洄游性鱼类;要么在淡水中没有随后的觅食,或者任何觅食都伴随着很少的体生长;主要的觅食和生长生物群落(海洋)与繁殖生物群落(淡水)不同。”
Myers defined catadromy as “diadromous fishes that spend most of their lives in freshwater before the migration of fully grown, adult fish to sea to reproduce; there is either no subsequent feeding at sea or any feeding is accompanied by little somatic growth; the principal feeding and growing biome (freshwater) differ from the reproductive biome (the sea).”
迈尔斯将降河洄游性定义为“在完全长大的成年鱼迁徙到海中繁殖之前,大部分生命时间在淡水中度过的洄游性鱼类;在海中要么没有随后的觅食,或者任何觅食都伴随着很少的体生长;主要的觅食和生长生物群落(淡水)与繁殖生物群落(海洋)不同。”
The term amphidromy was proposed by Myers to describe diadromous life histories where migrations between freshwater and seawater were not directly associated with reproduction.
两栖洄游性这个术语是由迈尔斯提出的,用来描述那些在淡水和海水之间的迁徙并不直接与繁殖相关联的洄游性生活史。
This difference in migration purpose (reproductive versus not reproductive) is the critical distinction between amphidromy and the other two types of diadromy.
迁徙目的的不同(繁殖性与非繁殖性)是两栖洄游性与其他两种洄游性类型之间的关键区别。
In addition, amphidromy is characterized by an obligate period of growth in an osmotic environment different from the hatching environment; Myers referred to this osmotic transition as “osmoregulatory migration.”
此外,两栖洄游性的特征是在与孵化环境不同的渗透环境中有一个必需的生长期;迈尔斯将这种渗透转变称为“渗透调节迁徙”。
Myers defined amphidromy as “diadromous fishes whose migration from freshwater to the sea, or vice- versa, is not for breeding but occurs regularly at some other definite stage of the life cycle.”
迈尔斯将两栖洄游性定义为“那些从淡水迁徙到海洋,或相反,但不是为了繁殖,而是在生命周期的某个其他特定阶段定期发生迁徙的洄游性鱼类。”
McDowall refined the definition as follows: “diadromous fishes in which there is a migration of larval fish to sea soon after hatching, followed by early feeding and growth at sea, and then a migration of small post-larval to juvenile fish from the sea back into freshwater; there is further, prolonged feeding in freshwater, during which most somatic growth from juvenile to adult stages occurs, as well as sexual maturation and reproduction; the principal feeding biome is the same as the reproductive biome (freshwater).”
麦克道尔对定义进行了如下完善:“在孵化后不久,幼鱼就迁徙到海中,随后在海中进行早期觅食和生长,然后是小型后期幼体到幼鱼从海洋迁徙回淡水;在淡水中还有进一步的、长时间的觅食,在此期间,大多数从幼鱼到成体阶段的体生长发生,以及性成熟和繁殖;主要的觅食生物群落与繁殖生物群落(淡水)相同。”
Unfortunately, both of these definitions are inadequate and problematic when used to describe the entire life cycle.
不幸的是,这两种定义在用来描述整个生命周期时都是不充分和有问题的。
Myers’ definition is accurate but simplistic, while McDowall’s definition only applies to specific growth stages, such as larva and post-larva.
迈尔斯的定义准确但过于简单,而麦克道尔的定义只适用于特定的生长阶段,如幼虫和后期幼虫。
See text S2 for the definition of oceanodromy.
有关海洋洄游性的定义,请参见补充文本S2。
Potamodromous fish are considered the most numerous migratory fish species in rivers worldwide (76, 77) and are the most common migratory fish species affected by dams.
河川洄游性鱼类被认为是全球河流中最丰富的迁徙性鱼类种群(76, 77),也是受大坝影响最普遍的迁徙性鱼类。
However, potamodromy is far less well studied than diadromy, and there is no further classification available to distinguish potamodromous phenotypes with complex life cycles (72).
然而,河川洄游性的研究远不如洄游性充分,也没有进一步的分类来区分具有复杂生命周期的河川洄游性表型(72)。
Therefore, we subdivided potamodromy into two types based on the spawning environment: riverine potamodromy and lacustrine potamodromy.
因此,我们根据产卵环境将河川洄游性细分为两种类型:河流性河川洄游性和湖泊性河川洄游性。
We defined the riverine type as potamodromous fishes that must migrate to the upper reaches of the river for reproduction until they complete reproduction in the riverine running water environment under specific conditions.
我们将河流类型定义为必须迁移到河流上游进行繁殖的河川洄游性鱼类,直到它们在特定条件下在河流流动水环境中完成繁殖。
Juveniles drift downstream with the current and enter lentic areas of the river or lakes to feed until their gonads reach a specific stage and begin their upriver migration for reproduction.
幼鱼随水流下游,并进入河流的滞水区或湖泊觅食,直到它们的性腺达到特定阶段,然后开始上游迁徙进行繁殖。
Most migratory fish species in the Yangtze River and other rivers in China exhibit riverine potamodromy, such as the four major carp species (Mylopharyngodon piceus, Ctenonpharyngodon idellus, Hypophthalmichthys molitrix, and Aristichthys nobilis).
长江和中国其他河流中的大多数迁徙性鱼类表现出河流性河川洄游性,如四大鲤鱼种(鳙、鲢、鳊和鲂)。
We also defined the lacustrine type as potamodromous fishes that must reproduce in the lake and usually lay eggs on aquatic plants.
我们还定义了湖泊类型为必须在湖泊中繁殖并通常在水生植物上产卵的河川洄游性鱼类。
At a specific growth stage, they regularly migrate to the lotic rivers and return to the lake after a period.
在特定的生长阶段,它们会定期迁移到流动的河流,并在一段时间后返回湖泊。
An example of fish species exhibiting lacustrine potamodromy is the roach (Rutilus rutilus) in Lake Krankesjon, a shallow lake in southern Sweden (78).
一个展示湖泊性河川洄游性的鱼类例子是瑞典南部克兰克湖(Lake Krankesjon)中的鲱鱼(Rutilus rutilus)(78)。
These lacustrine potamodromous fishes are typically characterized by partial migration behavior.
这些湖泊性河川洄游性鱼类通常表现为部分迁徙行为。
Six descriptive life cycle models of migratory fish
迁徙性鱼类的六个描述性生命周期模型
The life cycle of migratory fish includes migrations of various stages.
迁徙性鱼类的生命周期包括各个阶段的迁徙。
Previous studies of fish life cycles have often relied on the accumulation of survey data, which is limited by sample size, the inability to track juveniles, and the geographic limitations of fishery data.
以前的鱼类生命周期研究常常依赖于调查数据的积累,这些数据受到样本大小的限制、无法追踪幼鱼以及渔业数据的地理限制。
As a result, a well-rounded understanding of fish migrations may not be achieved, and erroneous inferences may be made.
结果可能是无法全面理解鱼类迁徙,可能会得出错误的推断。
Therefore, we suggest using migration patterns and typical cases to create life cycle models for the fish.
因此,我们建议使用迁徙模式和典型案例来为鱼类创建生命周期模型。
This will establish a life cycle framework that aligns with the migratory characteristics of the fish and avoids omitting key migration aspects based solely on survey data.
这将建立一个与鱼类迁徙特性相一致的生命周期框架,并避免仅基于调查数据而遗漏关键的迁徙方面。
We conducted case studies on the entire life cycles of typical fish species whose migration patterns were well known.
我们对那些迁徙模式已知的典型鱼种的整个生命周期进行了案例研究。
We then generalized six descriptive life cycle models of migratory fish corresponding to six migration types/subtypes of diadromy and potamodromy, excluding oceanodromy, which is not relevant to dams.
然后,我们概括了六个迁徙性鱼类的描述性生命周期模型,这些模型对应于洄游性和河川洄游性的六种迁徙类型/亚型,不包括与大坝无关的海洋洄游性。
Here, we show three life cycle models of anadromous, freshwater amphidromous, and riverine potamodromous fishes related to this study.
在这里,我们展示了与本研究相关的溯河洄游性、淡水两栖洄游性和河流性河川洄游性鱼类的三个生命周期模型。
See text S2 for the other three models.
有关其他三个模型的详细信息,请参见文本S2。
First, the life cycle model of anadromous fish involves diadromous fish that migrate between freshwater and saltwater environments, spending most of their lives in the sea or saltwater lakes for feeding.
首先,溯河洄游性鱼类的生命周期模型涉及在淡水和咸水环境之间迁徙的洄游性鱼类,它们大部分生命时间在海洋或咸水湖中觅食。
Adults that have reached a specific stage of gonadal development and are ready to breed begin to enter the river and migrate upstream, feeding little during this upriver migration.
达到特定性腺发育阶段并准备繁殖的成鱼开始进入河流并向上游迁徙,在这次上游迁徙过程中很少觅食。
Post-spawners of semelparous species die (e.g., salmonids), while post-spawners of iteroparous species migrate rapidly downriver to the sea for feeding.
单次繁殖物种的产卵后个体会死亡(例如,鲑科鱼类),而多次繁殖物种的产卵后个体则迅速向下游迁徙到海洋觅食。
After hatching, larvae and juveniles likewise migrate downstream to the sea or saltwater lakes, feeding and growing during their downriver migration and then undergoing osmoregulation in the estuary before ultimately entering the sea.
孵化后,幼虫和幼鱼同样向下游迁徙到海洋或咸水湖,在其下游迁徙过程中觅食和生长,然后在河口进行渗透调节,最终进入海洋。
Second, the life cycle model of the freshwater amphidromous fish involves diadromous fish that migrate between freshwater and saltwater environments, spending most of their lives in freshwater for feeding and reproduction.
其次,淡水两栖洄游性鱼类的生命周期模型涉及在淡水和咸水环境之间迁徙的洄游性鱼类,它们大部分生命时间在淡水中觅食和繁殖。
After hatching, larvae and juveniles must migrate downstream to the sea for early feeding and growth in saltwater until they reach a specific growth stage and complete osmoregulation.
孵化后,幼虫和幼鱼必须向下游迁徙到海洋,在咸水中进行早期觅食和生长,直到它们达到特定的生长阶段并完成渗透调节。
At this point, the fish return from the sea to the river for further feeding, prolonged upriver migration, somatic growth (e.g., from subadult to adult and spawner), sexual maturation, and ultimately reproduction in the river.
在这一点上,鱼从海洋返回河流进行进一步的觅食、长时间的上游迁徙、体生长(例如,从亚成体到成体和产卵者)、性成熟,最终在河流中繁殖。
After reproduction, post-spawners disperse downstream to feed, mostly in freshwater, until their gonads are close to maturity and they begin the upriver migration to reproduce again.
繁殖后,产卵后的个体分散向下游觅食,大部分在淡水中,直到它们的性腺接近成熟,它们开始向上游迁徙再次繁殖。
Last, the life cycle model of the riverine potamodromous fish involves potamodromous fish that migrate within freshwater and spend most of their lives in the lentic river areas or lakes for feeding.
最后,河流性河川洄游性鱼类的生命周期模型涉及在淡水中迁徙的河川洄游性鱼类,它们大部分生命时间在河流滞水区或湖泊中觅食。
Sexually mature adults migrate to the upper reaches of the river for reproduction until they complete reproduction in the riverine running water environment under specific conditions.
性成熟的成鱼迁移到河流上游进行繁殖,直到它们在特定条件下在河流流动水环境中完成繁殖。
They lay adhesive and demersal eggs or floating eggs.
它们产下粘性沉底卵或浮性卵。
The eggs/larvae drift with the current, or juveniles migrate downstream, and they enter river lentic areas or lakes to feed and to develop into juveniles or subadults.
卵/幼虫随水流漂流,或幼鱼向下游迁徙,它们进入河流滞水区或湖泊觅食并发育成幼鱼或亚成体。
Until their gonads reach a specific stage, they begin the prolonged upriver migration for reproduction.
直到它们的性腺达到特定阶段,它们开始长时间的上游迁徙进行繁殖。
After reproduction, post-spawners migrate downstream to river lentic areas or lakes to feed until their gonads are close to maturity and they begin the upriver migration to reproduce again.
繁殖后,产卵后的个体向下游迁移到河流滞水区或湖泊觅食,直到它们的性腺接近成熟,它们开始向上游迁徙再次繁殖。
A procedure for determining migratory fish life cycle
确定迁徙性鱼类生命周期的程序
We propose a procedure for determining the entire life cycle of a migratory fish species by combining a life cycle model with sporadic limited data.
我们提出了一种通过将生命周期模型与零星有限数据相结合来确定迁徙性鱼类整个生命周期的程序。
First, we use the three-tiered taxonomic system to confirm the migration pattern of a given fish and then identify its corresponding life cycle model.
首先,我们使用三层级分类系统来确认给定鱼类的迁徙模式,然后确定其相应的生命周期模型。
Second, we collect historical catch and survey data at various growth stages and verify the reliability of these data.
其次,我们在不同生长阶段收集历史捕捞和调查数据,并验证这些数据的可靠性。
We determine the spawning, feeding, and overwintering areas of the fish and the upper and lower limits of the migration path by collecting data on the reproduction, distribution, age structure, and abundance of the fish.
我们通过收集有关鱼类繁殖、分布、年龄结构和数量的数据,确定鱼类的产卵、觅食和越冬区域以及迁徙路径的上下限。
This information is gathered from literature and field interviews with local fishermen.
这些信息是从文献和与当地渔民的实地访谈中收集的。
Third, we use the available data to specify the details of the bidirectional migration, including the onset times of juvenile downstream migration and adult (or subadult) reproductive migration.
第三,我们使用可用数据来详细说明双向迁徙的细节,包括幼鱼下游迁徙的开始时间和成体(或亚成体)繁殖迁徙的开始时间。
We also identify endogenous factors, such as gonadal development, and exogenous factors, such as water temperature or hydrological variations that trigger a shift in the bidirectional migrations.
我们还识别内源因素,如性腺发育,以及外源因素,如水温或水文变化,这些因素触发双向迁徙的转变。
Last, we will obtain a general understanding of the entire life cycle of the species.
最后,我们将获得对物种整个生命周期的总体理解。
A simplified model of dam barrier effects
大坝障碍效应的简化模型
The alteration of fish resources involves the transition of generations and the progression of age groups characterized by varied natural mortality rates and fishing mortality rates.
鱼类资源的改变涉及代际转换和年龄组的进展,这些年龄组的特点是不同的自然死亡率和捕捞死亡率。
Age-structured population models that track dynamic changes in females and males in each age group can be used to quantify interannual variations in age distributions and population structure.
追踪每个年龄组中雌性和雄性动态变化的年龄结构化种群模型可以用来量化年龄分布和种群结构的年际变化。
Fish breeding behavior and population size usually fluctuate depending on environmental factors, such as river hydrological change (79).
鱼类的繁殖行为和种群大小通常会根据环境因素(如河流水文变化)而波动(79)。
However, barring direct human interference or drastic changes in the natural environment, the age structures of fish, particularly spawners, are relatively stable (30).
然而,在没有直接人为干预或自然环境发生剧烈变化的情况下,鱼类的年龄结构,特别是产卵者的年龄结构,相对稳定(30)。
The effect of a given dam on the breeding activity and population size of a migratory fish depends on both dam attributes (e.g., its location, flow regulation capacity, inundation range, and impoundment time) and fish attributes (e.g., migration pattern and spatiotemporal distribution in the river).
一个特定大坝对迁徙性鱼类的繁殖活动和种群大小的影响取决于大坝属性(例如,其位置、流量调节能力、淹没范围和蓄水时间)和鱼类属性(例如,迁徙模式和在河流中的时空分布)。
To quantify the dam barrier effect on population fragmentation, we derived a universal population model with age-specific survival rates and valid stock ratios.
为了量化大坝障碍对种群破碎的影响,我们推导出了一个具有年龄特定存活率和有效种群比例的通用种群模型。
Solving the universal model for a specific fish species requires a substantial amount of biological and ecological data, including survival rates and valid stock ratios for all age cohorts.
解决特定鱼类物种的通用模型需要大量的生物和生态数据,包括所有年龄组的生存率和有效种群比例。
However, obtaining these data can be challenging (text S3).
然而,获取这些数据可能具有挑战性(文本S3)。
In the absence of sufficient data, we propose a simplified mathematical model of the dam barrier effect that requires less data (text S3): Nt = N0 ∙ β^(t−T), where T is the first year of a given stock decline caused by dams, indicating a time lag between dam construction and a given stock decline; t is the number of years since time T; N0 is the initial number of individuals in a given stock, such as the spawning stock, the recruitment stock, or the total population; Nt is the number of individuals in a given stock in the year t; and β is the dimensionless DIC (0 ≤ β ≤ 1).
在缺乏足够数据的情况下,我们提出了一个简化的大坝障碍效应数学模型,该模型需要较少的数据(文本S3):Nt = N0 ∙ β^(t−T),其中T是由大坝引起的特定种群下降的第一年,表示大坝建设与特定种群下降之间的时间滞后;t是自时间T以来的年数;N0是给定种群(如产卵群体、补充群体或总人口)的初始个体数;Nt是年份t中给定种群的个体数;β是无量纲的大坝影响系数(DIC)(0 ≤ β ≤ 1)。
The DIC is an integrated coefficient that reflects the dam barrier effect on the survival rate and the proportion of valid stock.
DIC是一个综合系数,反映了大坝障碍对存活率和有效种群比例的影响。
The simplified model offers a valuable analytical approach for studying the barrier effects of dams on data-poor migratory fish populations.
简化模型为研究大坝对数据匮乏的迁徙性鱼类种群的障碍效应提供了宝贵的分析方法。
Figure S6 illustrates the impact of DIC (β) on the relative population sizes (N/N0) at T = 0.
图S6展示了DIC(β)在T = 0时对相对种群大小(N/N0)的影响。
The results indicate that if β is less than 0.6, then the population size will become EX in less than a decade.
结果表明,如果β小于0.6,那么种群大小将在不到十年的时间内变得灭绝(EX)。
Conversely, if β is greater than 0.8, the population size can be sustained for over 20 years.
相反,如果β大于0.8,种群大小可以维持超过20年。
Estimation methods for the DIC
DIC的估算方法
The key to using the simplified model is the estimation of the DIC, which depends on the strength of the dam barrier effect and the life history plasticity of the migratory fish.
使用简化模型的关键是DIC的估算,这取决于大坝障碍效应的强度和迁徙性鱼类的生活史可塑性。
A larger value of β indicates a higher degree of life history plasticity and greater adaptability of the migratory fish to the dam barrier effect, implying a slower rate of population decline and vice versa.
β值较大表示生活史可塑性较高,迁徙性鱼类对大坝障碍效应的适应性较强,意味着种群下降速率较慢,反之亦然。
To estimate the key parameter, we recommend four methods based on the data status: the catch method, the abundance method, the valid stock method, and the analogy method.
为了估算关键参数,我们基于数据状况推荐四种方法:捕捞法、丰富度法、有效种群法和类比法。
The DICs may vary for a given fish depending on its life stage (e.g., spawner or juvenile) and spatial location (e.g., upstream or downstream of the dam).
对于特定的鱼,DIC值可能会因其生活阶段(例如,产卵者或幼鱼)和空间位置(例如,大坝上游或下游)而异。
Therefore, separate estimates must be made for each specific stock and its corresponding spatial location.
因此,必须为每个特定的种群及其相应的空间位置进行单独估算。
To ensure accuracy, a combined approach may be necessary due to the different attributes of the collected data.
为了确保准确性,可能需要综合方法,因为收集到的数据具有不同的属性。
However, assessing the data availability remains a challenge when using these methods.
然而,在使用这些方法时,评估数据可用性仍然是一个挑战。
It is important to consider the influence of factors other than dams on the data, which may be small but cannot be entirely ignored.
重要的是要考虑除大坝以外的其他因素对数据的影响,这些影响可能很小,但不能完全忽视。
The catch method is an estimation approach based on catch data.
捕捞法是一种基于捕捞数据的估算方法。
It assumes that the multi-year catch data in the fixed river section, under the same fishing conditions after dam construction, reflect the changes in population size.
它假设在大坝建成后,固定河段多年捕捞数据在相同捕捞条件下反映了种群大小的变化。
Therefore, the simplified model can be used to perform regression analysis on the data and obtain the DIC.
因此,可以使用简化模型对数据进行回归分析并获得DIC。
In this study, we applied this method to specific stocks of Chinese paddlefish, Chinese sucker, and C. guichenoti.
在本研究中,我们将这种方法应用于中华匙吻鲟、中华鲴和鯮(Coreius guichenoti)的特定种群。
The abundance method is an estimation approach based on population or stock size results from various years under the influence of dams.
丰富度法是一种基于大坝影响下多年人口或种群大小结果的估算方法。
It is usually applied to well-studied fish.
它通常应用于研究充分的鱼类。
These abundance data have been estimated in previous studies and can be used to estimate DICs for a given fish.
这些丰富度数据已在以前的研究中估算过,可以用来估算给定鱼类的DIC。
In this study, we applied this method to the Chinese sturgeon and the specific stock of C. guichenoti.
在本研究中,我们将这种方法应用于中华鲟和特定种群的鯮(Coreius guichenoti)。
The valid stock method is an estimation approach based on valid stocks.
有效种群法是一种基于有效种群的估算方法。
By analyzing the entire life cycle of fish, as well as fishing and survey data, we can estimate the distribution ranges and patterns of spawning and recruitment stocks along the migration route before dam construction.
通过分析鱼类的整个生命周期以及捕捞和调查数据,我们可以估算大坝建设前沿迁徙路线的产卵群体和补充群体的分布范围和模式。
On the basis of the information about the dam, we can estimate the proportion of invalid stocks in the spawning and recruitment stocks after dam construction.
根据大坝的信息,我们可以估算大坝建设后产卵群体和补充群体中无效种群的比例。
This allows us to obtain the DIC of the specific stock, which represents the proportion of the valid stock.
这使我们能够获得特定种群的DIC,它代表了有效种群的比例。
In this study, we applied this method to specific stocks of Yangtze sturgeon and C. guichenoti.
在本研究中,我们将这种方法应用于长江鲟和鯮(Coreius guichenoti)的特定种群。
The analogy method is an estimation approach based on a database containing numerous case studies.
类比法是一种基于包含众多案例研究的数据库的估算方法。
In the absence of available data, we searched for similar cases of other fish species to estimate the DIC for a given fish species.
在没有可用数据的情况下,我们寻找其他鱼类物种的类似案例来估算给定鱼类物种的DIC。
On the basis of the case studies in this study, we used the DIC of Chinese paddlefish by analogy to estimate the DIC (β = 0.53) of C. guichenoti, which was consistent with the result of the catch method (0.54).
根据本研究的案例研究,我们通过类比使用中华匙吻鲟的DIC来估算鯮(Coreius guichenoti)的DIC(β = 0.53),这与捕捞法的结果(0.54)一致。
Verification of the simplified model
简化模型的验证
To assess the accuracy and applicability of the simplified model, we compared our results with the capacity-gonad-confined population model (CGPM) for the Chinese sturgeon, which is a highly precise model based on age-structured functions (30).
为了评估简化模型的准确性和适用性,我们将结果与中国鲟的容量-性腺限制种群模型(CGPM)进行了比较,CGPM是一种基于年龄结构函数的高精度模型(30)。
Figure S7 (C and D) shows that the results of the simplified model were slightly lower than those of the CGPM in the early stages of dam operation.
图S7(C和D)显示,在大坝运行的早期阶段,简化模型的结果略低于CGPM的结果。
Over time, however, the discrepancy between the two model estimates decreased and the estimated species extinction times became more consistent.
然而,随着时间的推移,两种模型估计之间的差异减小,估算的物种灭绝时间变得更加一致。
Therefore, we concluded that the simplified model provides a valuable estimate of dam effects on population size, even in cases where detailed biological parameters are not available.
因此,我们得出结论,简化模型即使在没有详细的生物参数的情况下,也能提供对大坝对种群大小影响的宝贵估计。
The predictions of our model were also verified by fishing and survey data, including the extinction time of the Chinese paddlefish and Yangtze sturgeon (18, 51).
我们模型的预测还通过捕捞和调查数据进行了验证,包括中华匙吻鲟和长江鲟的灭绝时间(18, 51)。
This confirmation demonstrates that the simplified model can accurately forecast the impact of the dam on the migratory fish population with limited data.
这一确认表明,简化模型可以准确预测大坝对迁徙性鱼类种群的影响,即使数据有限。
See text S4 for details.
有关详细信息,请参见文本S4。
Limitations of this study
本研究的局限性
This study has certain limitations, such as the need for larger sample sizes of fish to improve the accuracy of the precision of fish life cycle models.
本研究存在某些局限性,例如需要更大的鱼类样本量来提高鱼类生命周期模型的准确性和精确度。
It is also necessary to refine the parameter estimation methods for the simplified model to accommodate various scenarios of data-limited fish.
还需要完善简化模型的参数估算方法,以适应数据有限的鱼类的各种情况。
In addition, further refinement and quantification of the invalid stock is needed to establish a more robust correlation between the invalid stock and the DIC.
此外,需要进一步改进和量化无效种群,以建立无效种群与DIC之间更稳健的相关性。
Supplementary Materials
补充材料
This PDF file includes: Supplementary text S1 to S5 Figs. S1 to S13 tables S1 to S7 Legends for movies S1 to S5 References
此PDF文件包括:补充文本S1至S5、图S1至S13、表格S1至S7、电影S1至S5的图例、参考文献。
Other Supplementary Material for this manuscript includes the following:
本文的其他补充材料包括以下内容:Movies S1 to S5
Acknowledgments:
We thank H. Zhang at the Yangtze River Fisheries Research Institute for sharing the Chinese sturgeon bycatch data.
我们感谢长江水产研究所的张慧女士分享中华鲟的兼捕数据。
We extend our gratitude to L. Wang at the China Institute of Water Resources and Hydropower Research, C. Li at the Chongqing Academy of Environmental Sciences, and T. Mu (former Head of Yibin Fishery Administration Office in Sichuan Province) for valuable assistance in conducting field surveys of the spawning grounds and stocks of the five fish species in 2017 and 2023.
我们向中国水利水电科学研究院的王磊先生、重庆环境科学院的李春先生以及四川省宜宾渔政管理办公室前主任牟涛先生表示感谢,感谢他们在2017年和2023年对五个鱼种的产卵场和种群进行实地调查时提供的宝贵帮助。
We are also grateful to X. Ke (former director of Sichuan Aquatic Research Institute, Sichuan Province) and other anonymous experts who were interviewed by us in 2017 and provided technical details of their involvement in the Gd-FRP.
我们同样感谢四川省水产研究所前所长柯贤章先生以及我们在2017年采访的提供其参与葛洲坝鱼类救援计划(Gd-FRP)技术细节的其他匿名专家。
Funding: this research was supported by the National Natural Science Foundation of China (52079148).
资金支持:本研究由中国国家自然科学基金(52079148)资助。
Author contributions: conceptualization: Z.H. Methodology: Z.H. Investigation: Z.H. and H.L. Visualization: Z.H. and H.L. Funding acquisition: Z.H. Project administration: Z.H. Supervision: Z.H. Writing—original draft: Z.H. and H.L. Writing—review and editing: Z.H.
作者贡献:概念化:Z.H. 方法论:Z.H. 调查:Z.H. 和 H.L. 可视化:Z.H. 和 H.L. 资金获取:Z.H. 项目管理:Z.H. 监督:Z.H. 写作—原稿撰写:Z.H. 和 H.L. 写作—审稿和编辑:Z.H.
Competing interests: the authors declare that they have no competing interests.
竞争利益:作者声明他们没有竞争利益。
Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials.
数据和材料可用性:评估论文中结论所需的所有数据均在论文和/或补充材料中提供

